

Dendritic cell podosomes are protrusive and invade the extracellular matrix using metalloproteinase MMP-14
- PMID: 20356925
- PMCID: PMC2858019
- DOI: 10.1242/jcs.056515
Dendritic cell podosomes are protrusive and invade the extracellular matrix using metalloproteinase MMP-14
Abstract
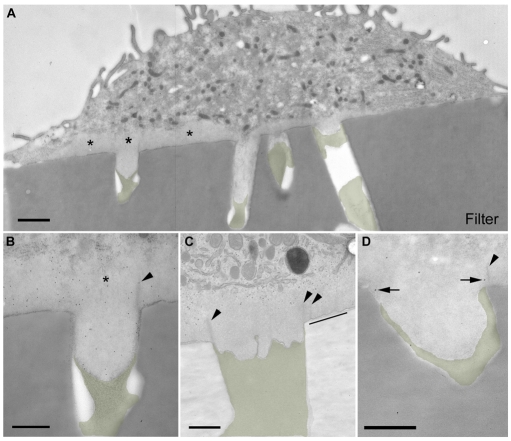
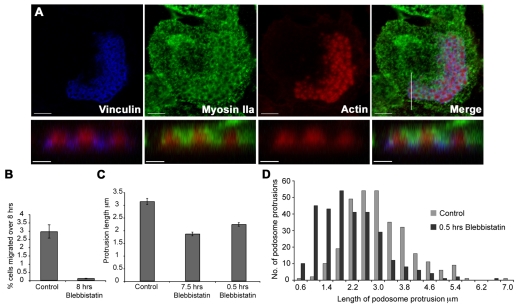
Podosomes are spot-like actin-rich structures formed at the ventral surface of monocytic and haematopoietic cells. Podosomes degrade extracellular matrix and are proposed to be involved in cell migration. A key question is whether podosomes form protrusions similar to the invadopodia of cancer cells. We characterised podosomes of immature dendritic cells using electron microscopy combined with both conventional and novel high-resolution structured illumination light microscopy. Dendritic cell podosomes are composed of actin foci surrounded by a specialised ring region that is rich in material containing paxillin. We found that podosomes were preferential sites for protrusion into polycarbonate filters impregnated with crosslinked gelatin, degrading up to 2 micrometers of matrix in 24 hours. Podosome-associated uptake of colloidal gold-labelled gelatin matrix appeared to occur via large phagosome-like structures or narrow tubular invaginations. The motor protein myosin-II was excluded from ring or core regions but was concentrated around them and the myosin-II inhibitor Blebbistatin reduced the length of podosome protrusions. Finally, we found that degradation, protrusion and endocytosis in this system are dependent on the matrix metalloproteinase MMP-14. We propose that podosomes mediate migration of dendritic cells through tissues by means of myosin-II-dependent protrusion coupled to MMP-14-dependent degradation and endocytosis.
Figures






References
-
- Akisaka T., Yoshida H., Suzuki R., Takama K. (2008). Adhesion structures and their cytoskeleton-membrane interactions at podosomes of osteoclasts in culture. Cell Tissue Res. 331, 625-641 - PubMed
-
- Baldassarre M., Ayala I., Beznoussenko G., Giacchetti G., Machesky L. M., Luini A., Buccione R. (2006). Actin dynamics at sites of extracellular matrix degradation. Eur. J. Cell Biol. 85, 1217-1231 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
