

CLIC5 mutant mice are resistant to diet-induced obesity and exhibit gastric hemorrhaging and increased susceptibility to torpor
- PMID: 20357015
- PMCID: PMC2886706
- DOI: 10.1152/ajpregu.00849.2009
CLIC5 mutant mice are resistant to diet-induced obesity and exhibit gastric hemorrhaging and increased susceptibility to torpor
Abstract
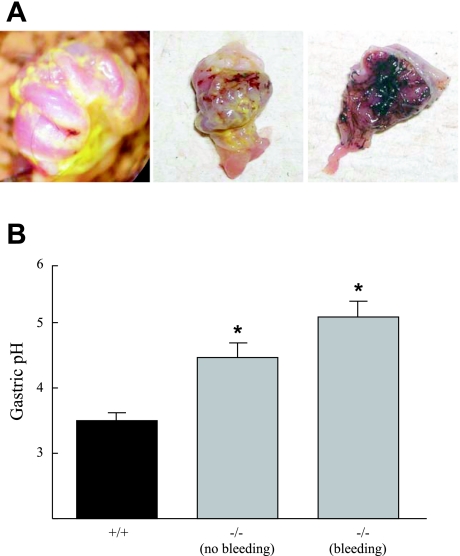
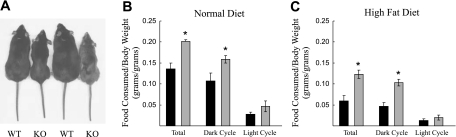
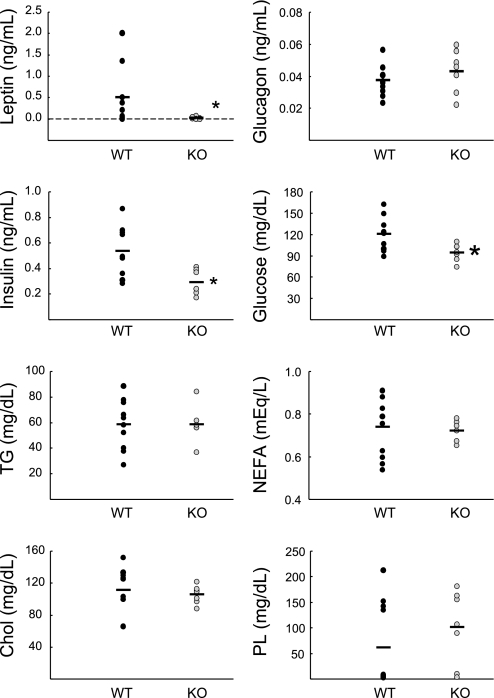
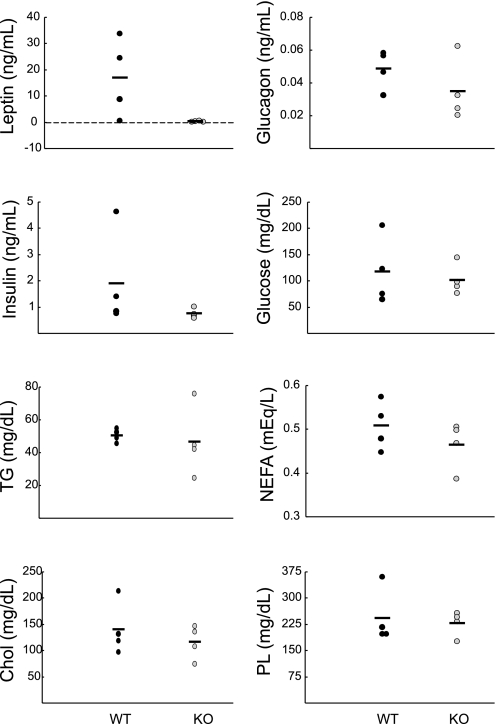
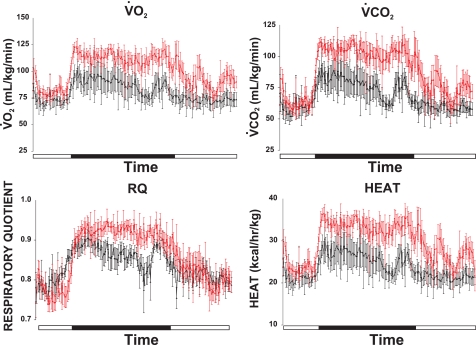
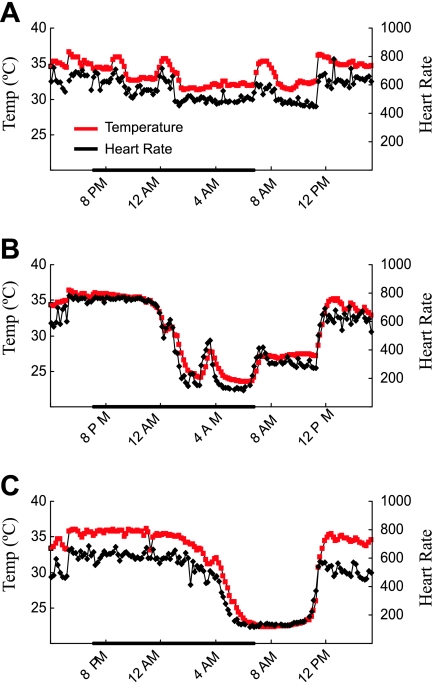
Chloride intracellular channel 5 (CLIC5) and other CLIC isoforms have been implicated in a number of biological processes, but their specific functions are poorly understood. The association of CLIC5 with ezrin and the actin cytoskeleton led us to test its possible involvement in gastric acid secretion. Clic5 mutant mice exhibited only a minor reduction in acid secretion, Clic5 mRNA was expressed at only low levels in stomach, and Clic5 mutant parietal cells were ultrastructurally normal, negating the hypothesis that CLIC5 plays a major role in acid secretion. However, the mutants exhibited gastric hemorrhaging in response to fasting, reduced monocytes and granulocytes suggestive of immune dysfunction, behavioral and social disorders suggestive of neurological dysfunction, and evidence of a previously unidentified metabolic defect. Wild-type and mutant mice were maintained on normal and high-fat diets; plasma levels of various hormones, glucose, and lipids were determined; and body composition was studied by quantitative magnetic resonance imaging. Clic5 mutants were lean, hyperphagic, and highly resistant to diet-induced obesity. Plasma insulin and glucose levels were reduced, and leptin levels were very low; however, plasma triglycerides, cholesterol, phospholipids, and fatty acids were normal. Indirect calorimetry revealed increased peripheral metabolism and greater reliance on carbohydrate metabolism. Because Clic5 mutants were unable to maintain energy reserves, they also exhibited increased susceptibility to fasting-induced torpor, as indicated by telemetric measurements showing episodes of reduced body temperature and heart rate. These data reveal a requirement for CLIC5 in the maintenance of normal systemic energy metabolism.
Figures


References
-
- Ashley RH. Challenging accepted ion channel biology: p64 and the CLIC family of putative intracellular anion channel proteins. Mol Membr Biol 20: 1–11, 2003 - PubMed
-
- Bennett BD, Solar GP, Yuan JQ, Mathias J, Thomas GR, Matthews W. A role for leptin and its cognate receptor in hematopoiesis. Curr Biol 6: 1170–1180, 1996 - PubMed
-
- Berry KL, Bulow HE, Hall DH, Hobert OA. C. elegans CLIC-like protein required for intracellular tube formation and maintenance. Science 302: 2134–2137, 2003 - PubMed
-
- Berryman M, Bruno J, Price J, Edwards JC. CLIC-5A functions as a chloride channel in vitro and associates with the cortical actin cytoskeleton in vitro and in vivo. J Biol Chem 279: 34794–34801, 2004 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases