

Discrimination of communication vocalizations by single neurons and groups of neurons in the auditory midbrain
- PMID: 20357062
- PMCID: PMC2888246
- DOI: 10.1152/jn.01131.2009
Discrimination of communication vocalizations by single neurons and groups of neurons in the auditory midbrain
Abstract
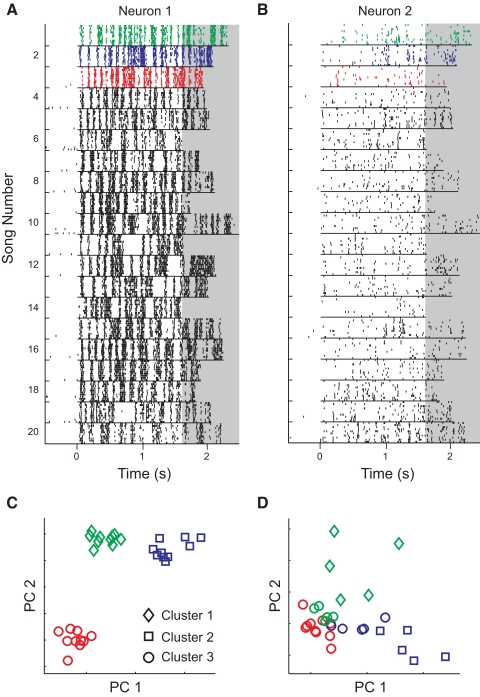
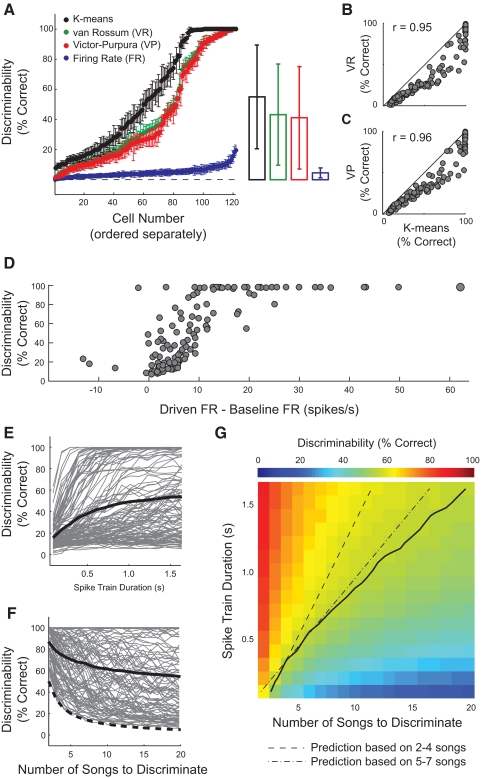
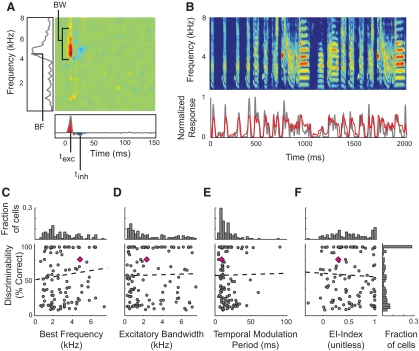
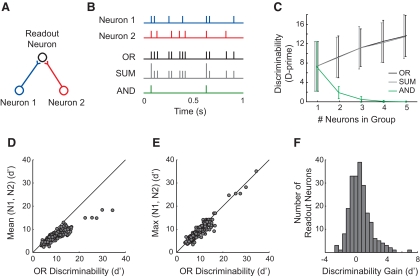
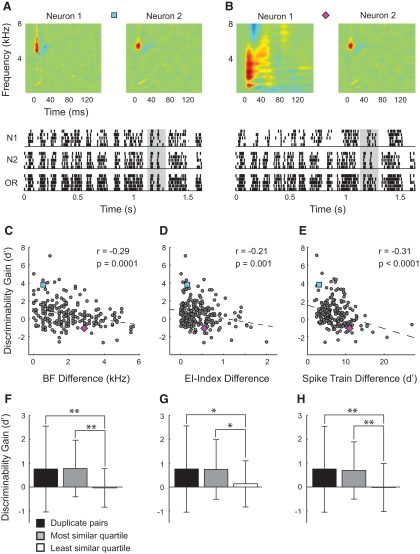
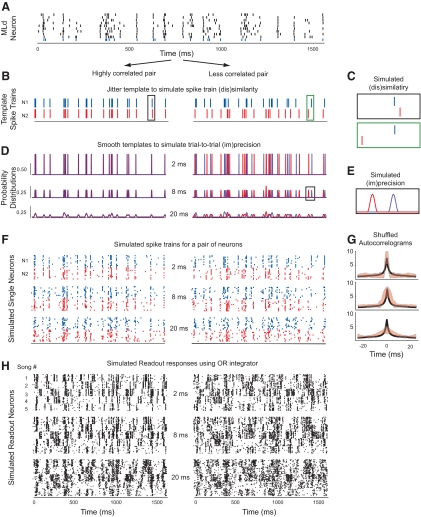
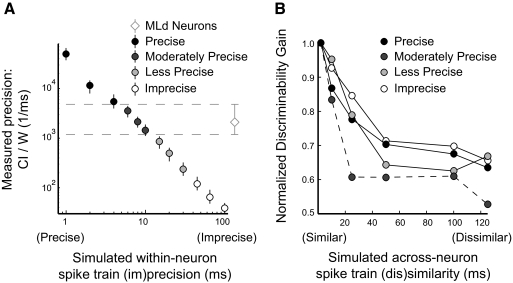
Many social animals including songbirds use communication vocalizations for individual recognition. The perception of vocalizations depends on the encoding of complex sounds by neurons in the ascending auditory system, each of which is tuned to a particular subset of acoustic features. Here, we examined how well the responses of single auditory neurons could be used to discriminate among bird songs and we compared discriminability to spectrotemporal tuning. We then used biologically realistic models of pooled neural responses to test whether the responses of groups of neurons discriminated among songs better than the responses of single neurons and whether discrimination by groups of neurons was related to spectrotemporal tuning and trial-to-trial response variability. The responses of single auditory midbrain neurons could be used to discriminate among vocalizations with a wide range of abilities, ranging from chance to 100%. The ability to discriminate among songs using single neuron responses was not correlated with spectrotemporal tuning. Pooling the responses of pairs of neurons generally led to better discrimination than the average of the two inputs and the most discriminating input. Pooling the responses of three to five single neurons continued to improve neural discrimination. The increase in discriminability was largest for groups of neurons with similar spectrotemporal tuning. Further, we found that groups of neurons with correlated spike trains achieved the largest gains in discriminability. We simulated neurons with varying levels of temporal precision and measured the discriminability of responses from single simulated neurons and groups of simulated neurons. Simulated neurons with biologically observed levels of temporal precision benefited more from pooling correlated inputs than did neurons with highly precise or imprecise spike trains. These findings suggest that pooling correlated neural responses with the levels of precision observed in the auditory midbrain increases neural discrimination of complex vocalizations.
Figures

Similar articles
-
Stimulus-dependent auditory tuning results in synchronous population coding of vocalizations in the songbird midbrain.J Neurosci. 2006 Mar 1;26(9):2499-512. doi: 10.1523/JNEUROSCI.3731-05.2006. J Neurosci. 2006. PMID: 16510728 Free PMC article.
-
Anesthetic state modulates excitability but not spectral tuning or neural discrimination in single auditory midbrain neurons.J Neurophysiol. 2011 Aug;106(2):500-14. doi: 10.1152/jn.01072.2010. Epub 2011 May 4. J Neurophysiol. 2011. PMID: 21543752 Free PMC article.
-
A spike-timing code for discriminating conspecific vocalizations in the thalamocortical system of anesthetized and awake guinea pigs.J Neurosci. 2009 Jan 14;29(2):334-50. doi: 10.1523/JNEUROSCI.3269-08.2009. J Neurosci. 2009. PMID: 19144834 Free PMC article.
-
How do auditory cortex neurons represent communication sounds?Hear Res. 2013 Nov;305:102-12. doi: 10.1016/j.heares.2013.03.011. Epub 2013 Apr 17. Hear Res. 2013. PMID: 23603138 Review.
-
Auditory processing of vocal sounds in birds.Curr Opin Neurobiol. 2006 Aug;16(4):400-7. doi: 10.1016/j.conb.2006.07.003. Epub 2006 Jul 13. Curr Opin Neurobiol. 2006. PMID: 16842993 Review.
Cited by
-
Functional architecture of the inferior colliculus revealed with voltage-sensitive dyes.Front Neural Circuits. 2013 Mar 20;7:41. doi: 10.3389/fncir.2013.00041. eCollection 2013. Front Neural Circuits. 2013. PMID: 23518906 Free PMC article.
-
Diverse cortical codes for scene segmentation in primate auditory cortex.J Neurophysiol. 2015 Apr 1;113(7):2934-52. doi: 10.1152/jn.01054.2014. Epub 2015 Feb 18. J Neurophysiol. 2015. PMID: 25695655 Free PMC article.
-
GABAA receptors contribute more to rate than temporal coding in the IC of awake mice.J Neurophysiol. 2020 Jan 1;123(1):134-148. doi: 10.1152/jn.00377.2019. Epub 2019 Nov 13. J Neurophysiol. 2020. PMID: 31721644 Free PMC article.
-
Auditory cortex shapes sound responses in the inferior colliculus.Elife. 2020 Jan 31;9:e51890. doi: 10.7554/eLife.51890. Elife. 2020. PMID: 32003747 Free PMC article.
-
Inhibition does not affect the timing code for vocalizations in the mouse auditory midbrain.Front Physiol. 2014 Apr 16;5:140. doi: 10.3389/fphys.2014.00140. eCollection 2014. Front Physiol. 2014. PMID: 24795640 Free PMC article.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources