

Retroviral reverse transcriptases
- PMID: 20358252
- PMCID: PMC11115783
- DOI: 10.1007/s00018-010-0346-2
Retroviral reverse transcriptases
Abstract
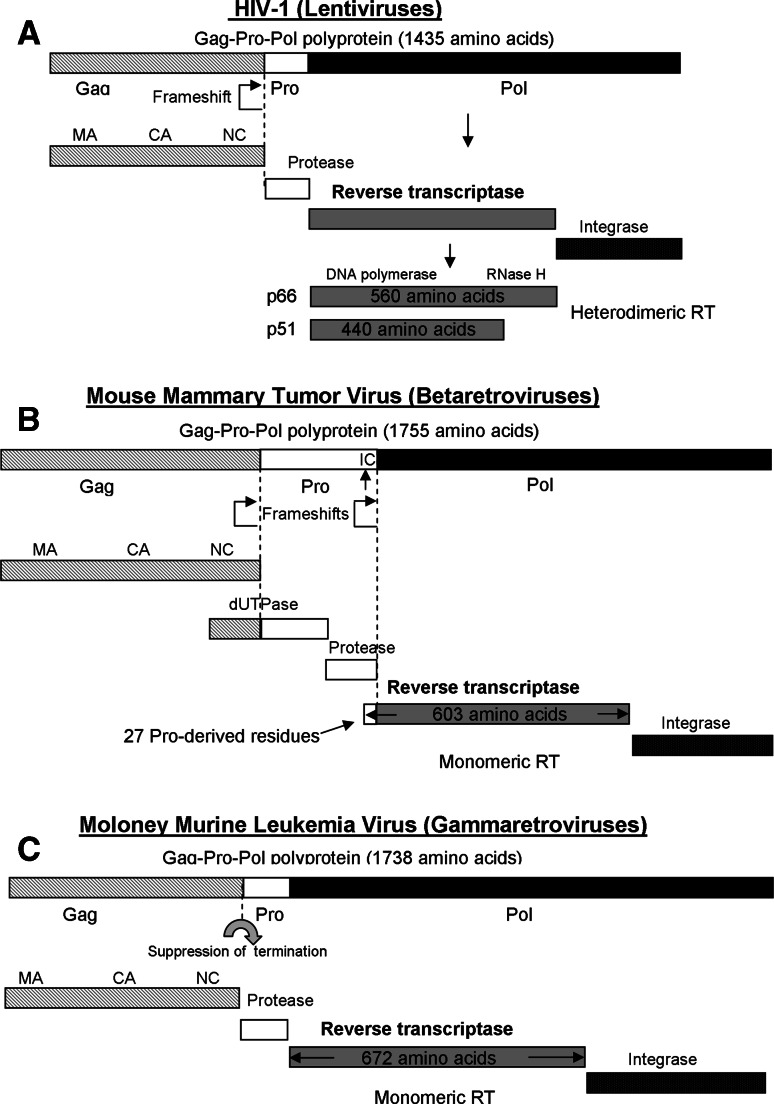
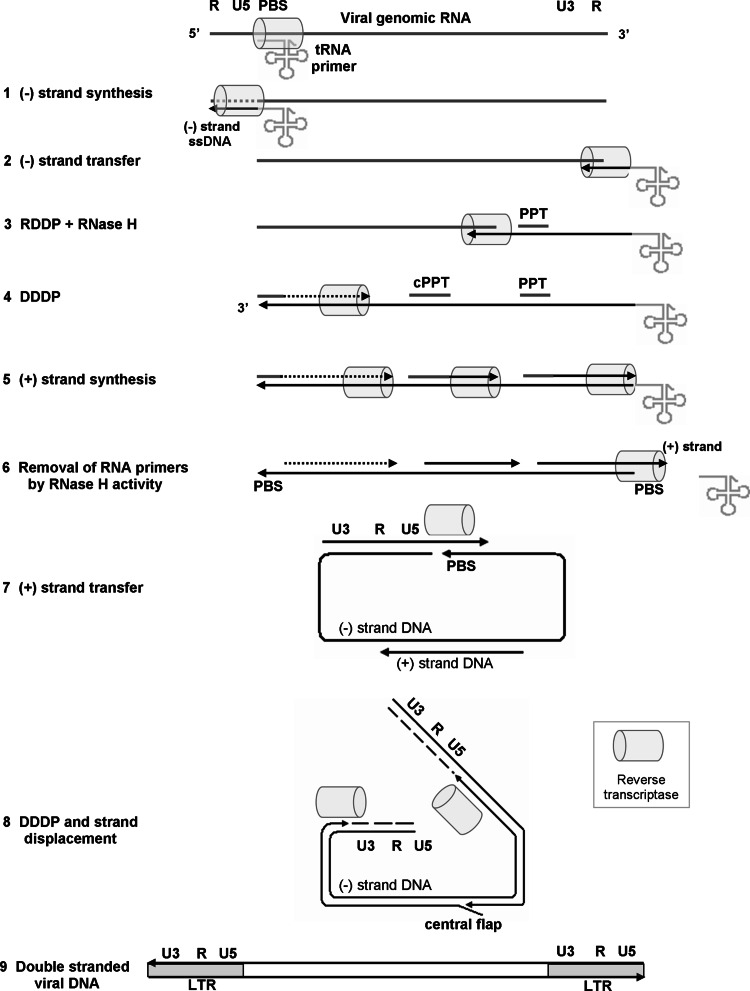
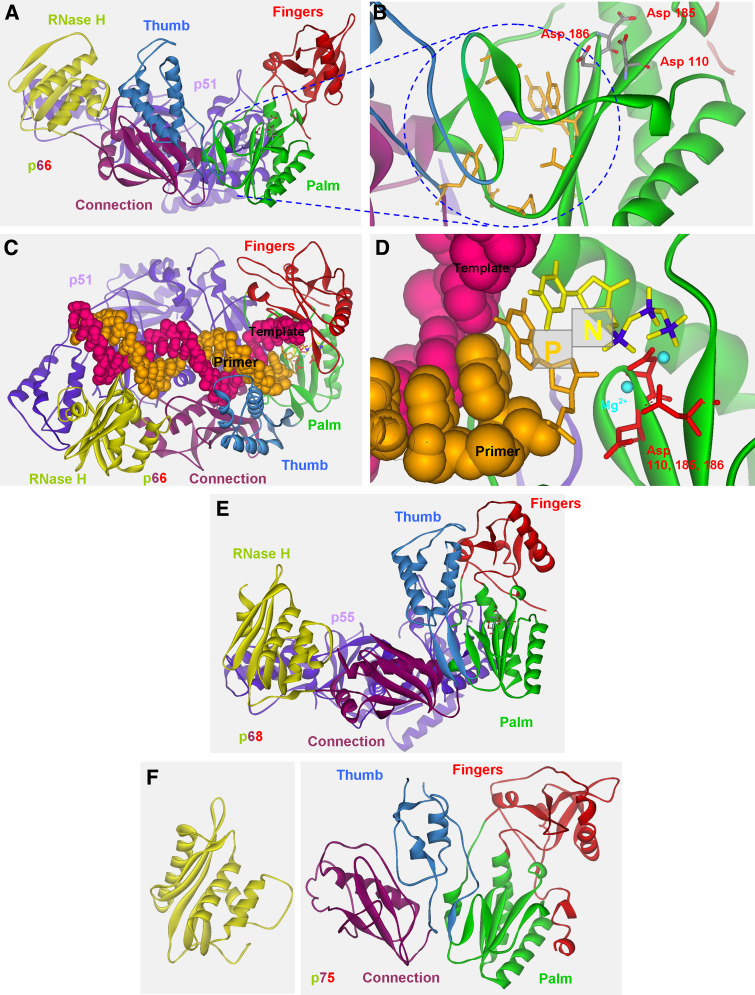
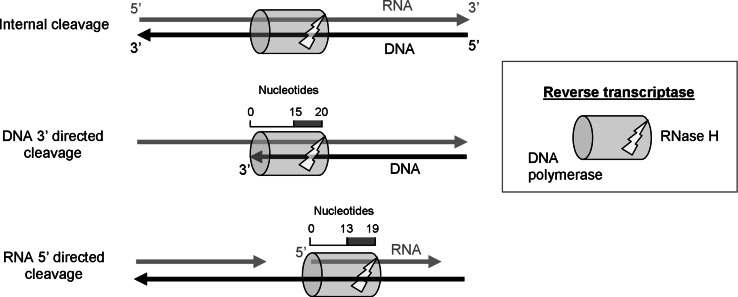
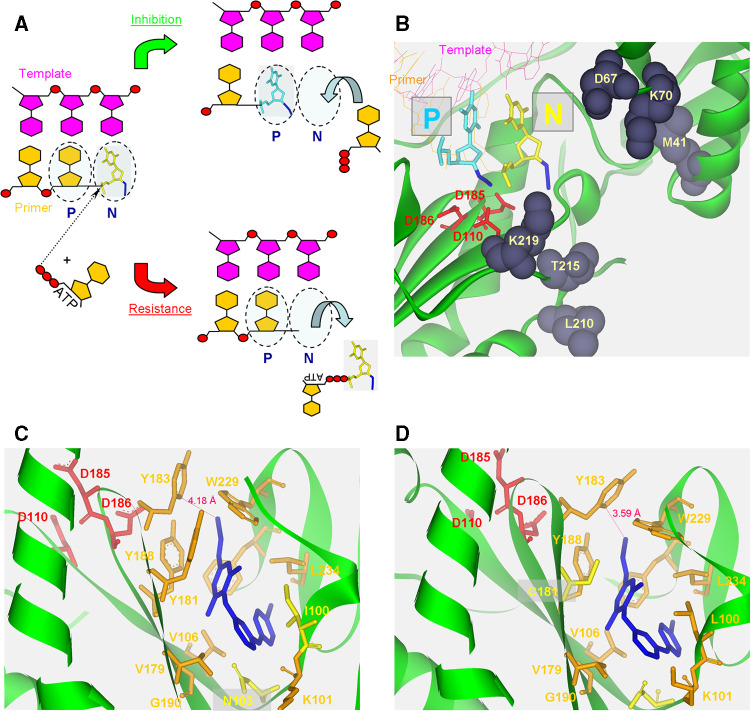
Reverse transcription is a critical step in the life cycle of all retroviruses and related retrotransposons. This complex process is performed exclusively by the retroviral reverse transcriptase (RT) enzyme that converts the viral single-stranded RNA into integration-competent double-stranded DNA. Although all RTs have similar catalytic activities, they significantly differ in several aspects of their catalytic properties, their structures and subunit composition. The RT of human immunodeficiency virus type-1 (HIV-1), the virus causing acquired immunodeficiency syndrome (AIDS), is a prime target for the development of antiretroviral drug therapy of HIV-1/AIDS carriers. Therefore, despite the fundamental contributions of other RTs to the understanding of RTs and retrovirology, most recent RT studies are related to HIV-1 RT. In this review we summarize the basic properties of different RTs. These include, among other topics, their structures, enzymatic activities, interactions with both viral and host proteins, RT inhibition and resistance to antiretroviral drugs.
Figures
References
-
- Coffin JM, Hughes SH, Varmus HE. Retroviruses. Cold Spring Harbor: Cold Spring Harbor Laboratory Press; 1997. - PubMed
-
- Skalka AM, Goff SP. Reverse transcriptase. Cold Spring Harbor: Cold Spring Harbor Laboratory Press; 1993. - PubMed
-
- Menendez-Arias L, Berkhout B (2008) Special issue on: retroviral reverse transcription. Virus Res 134 - PubMed
-
- Parniak MA (2004) Special issue: molecular biology of HIV. Int J Biochem Cell Biol 36
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
