

A gating charge transfer center in voltage sensors
- PMID: 20360102
- PMCID: PMC2869078
- DOI: 10.1126/science.1185954
A gating charge transfer center in voltage sensors
Abstract
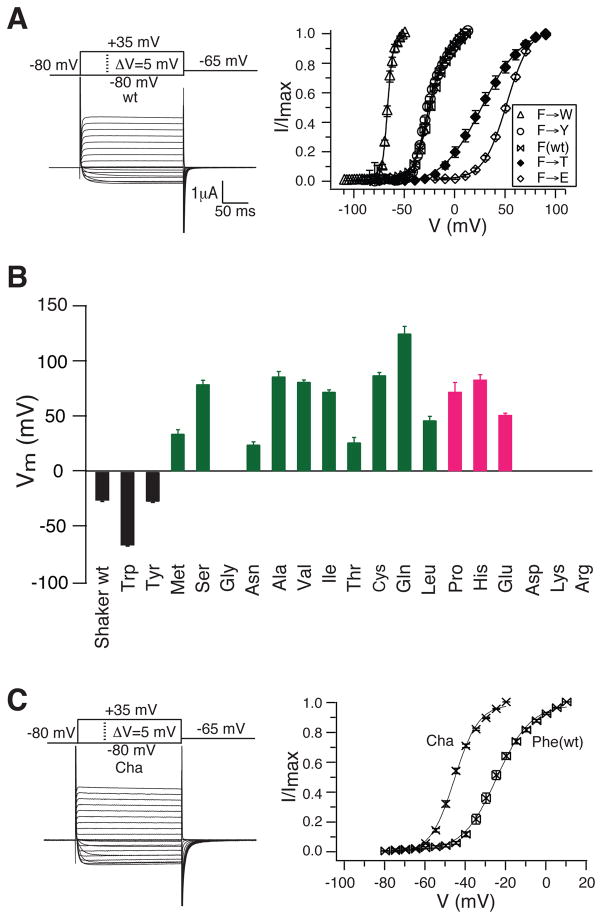
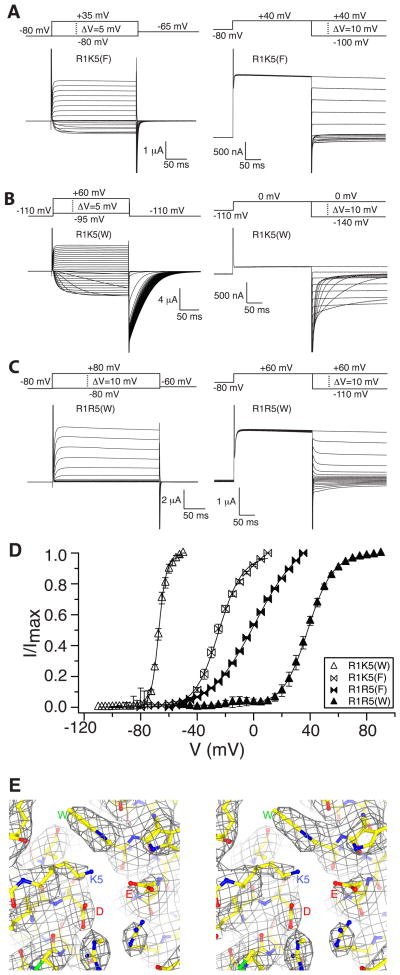
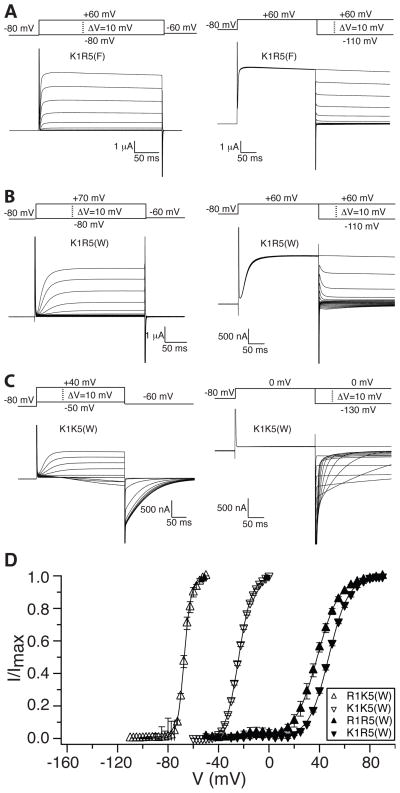
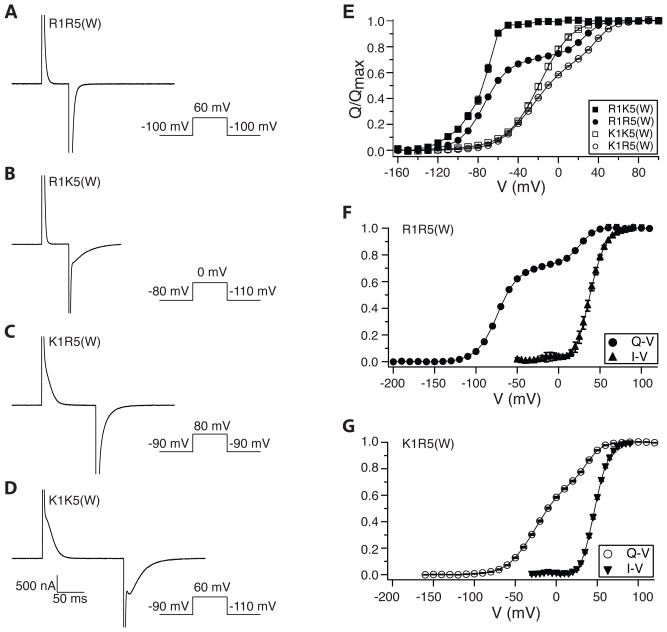
Voltage sensors regulate the conformations of voltage-dependent ion channels and enzymes. Their nearly switchlike response as a function of membrane voltage comes from the movement of positively charged amino acids, arginine or lysine, across the membrane field. We used mutations with natural and unnatural amino acids, electrophysiological recordings, and x-ray crystallography to identify a charge transfer center in voltage sensors that facilitates this movement. This center consists of a rigid cyclic "cap" and two negatively charged amino acids to interact with a positive charge. Specific mutations induce a preference for lysine relative to arginine. By placing lysine at specific locations, the voltage sensor can be stabilized in different conformations, which enables a dissection of voltage sensor movements and their relation to ion channel opening.
Figures


References
-
- Hille B. Ion Channels of Excitable Membranes. Sinauer Associates; Sunderland, MA: 2001.
-
- Okamura Y, Murata Y, Iwasaki H, Sasaki M. Tanpakushitsu Kakusan Koso. 2006 Jan;51:18. - PubMed
-
- Murata Y, Iwasaki H, Sasaki M, Inaba K, Okamura Y. Nature. 2005 Jun 30;435:1239. - PubMed
-
- Seoh SA, Sigg D, Papazian DM, Bezanilla F. Neuron. 1996 Jun;16:1159. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
