

The nuclear receptor tailless induces long-term neural stem cell expansion and brain tumor initiation
- PMID: 20360385
- PMCID: PMC2849125
- DOI: 10.1101/gad.560310
The nuclear receptor tailless induces long-term neural stem cell expansion and brain tumor initiation
Abstract
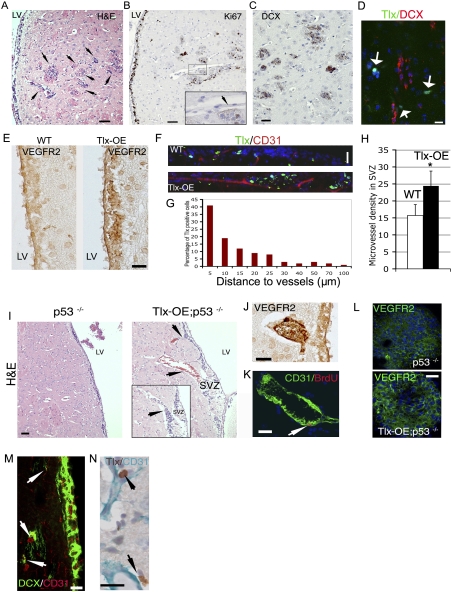
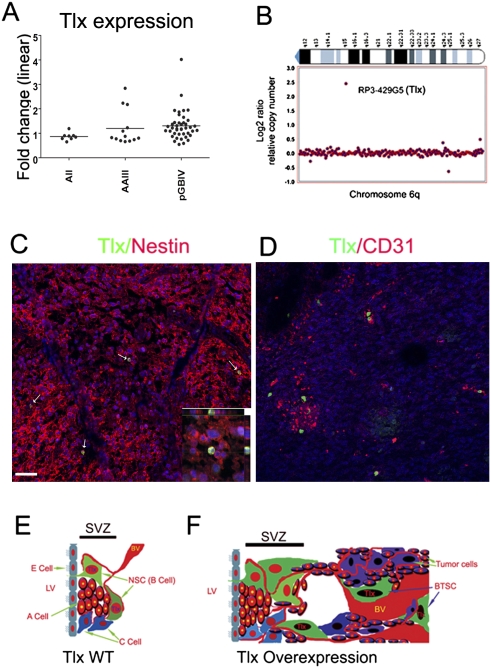
Malignant gliomas are the most common primary brain tumors, and are associated with frequent resistance to therapy as well as poor prognosis. Here we demonstrate that the nuclear receptor tailless (Tlx), which in the adult is expressed exclusively in astrocyte-like B cells of the subventricular zone, acts as a key regulator of neural stem cell (NSC) expansion and brain tumor initiation from NSCs. Overexpression of Tlx antagonizes age-dependent exhaustion of NSCs in mice and leads to migration of stem/progenitor cells from their natural niche. The increase of NSCs persists with age, and leads to efficient production of newborn neurons in aged brain tissues. These cells initiate the development of glioma-like lesions and gliomas. Glioma development is accelerated upon loss of the tumor suppressor p53. Tlx-induced NSC expansion and gliomagenesis are associated with increased angiogenesis, which allows for the migration and maintenance of brain tumor stem cells in the perivascular niche. We also demonstrate that Tlx transcripts are overexpressed in human primary glioblastomas in which Tlx expression is restricted to a subpopulation of nestin-positive perivascular tumor cells. Our study clearly demonstrates how NSCs contribute to brain tumorgenesis driven by a stem cell-specific transcription factor, thus providing novel insights into the histogenesis and molecular pathogenesis of primary brain tumors.
Figures
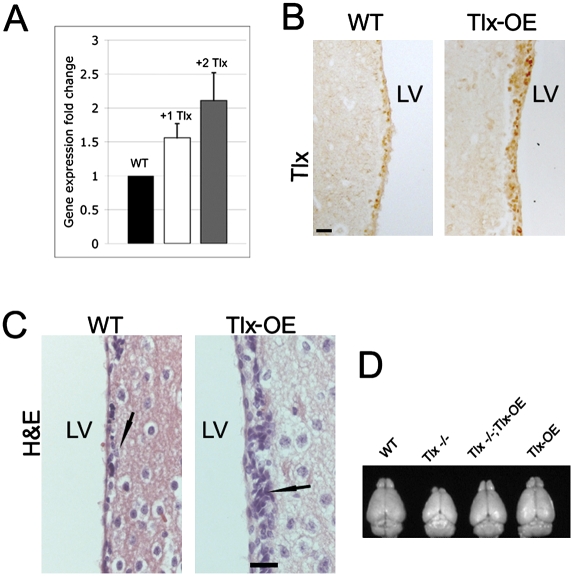
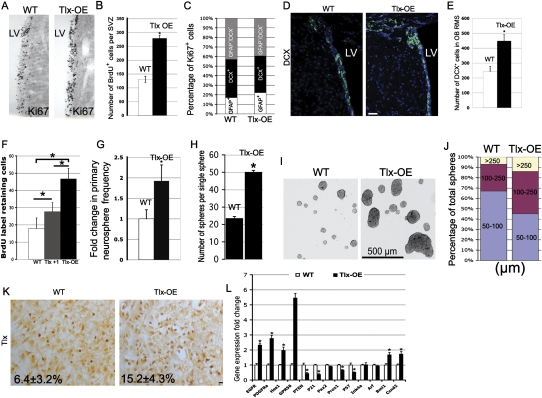
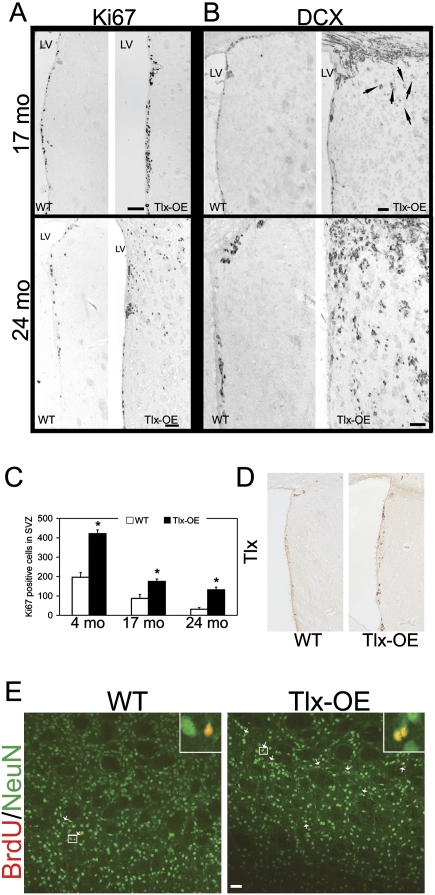
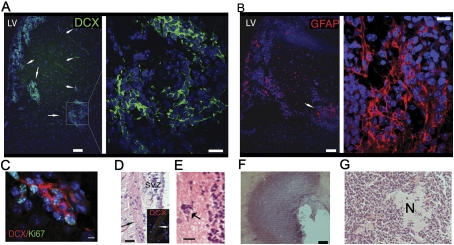

References
-
- Androutsellis-Theotokis A, Leker RR, Soldner F, Hoeppner DJ, Ravin R, Poser SW, Rueger MA, Bae SK, Kittappa R, McKay RD. Notch signalling regulates stem cell numbers in vitro and in vivo. Nature. 2006;442:823–826. - PubMed
-
- Antonchuk J, Sauvageau G, Humphries RK. HOXB4-induced expansion of adult hematopoietic stem cells ex vivo. Cell. 2002;109:39–45. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous