

The nuclear hormone receptor Coup-TFII is required for the initiation and early maintenance of Prox1 expression in lymphatic endothelial cells
- PMID: 20360386
- PMCID: PMC2849126
- DOI: 10.1101/gad.1859310
The nuclear hormone receptor Coup-TFII is required for the initiation and early maintenance of Prox1 expression in lymphatic endothelial cells
Abstract
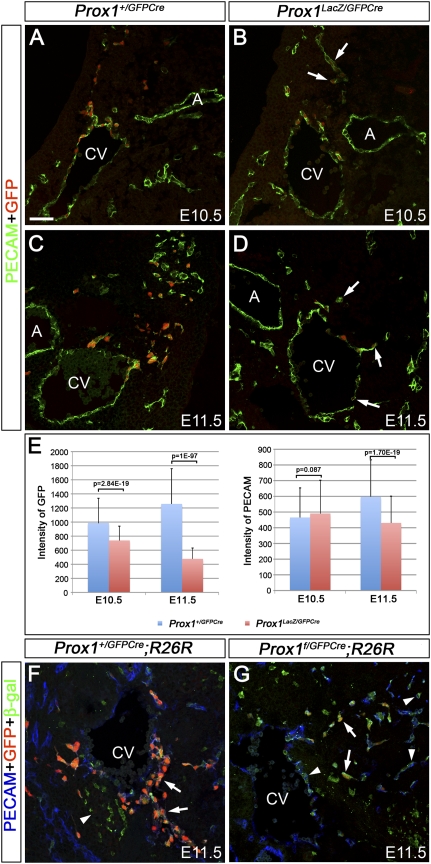
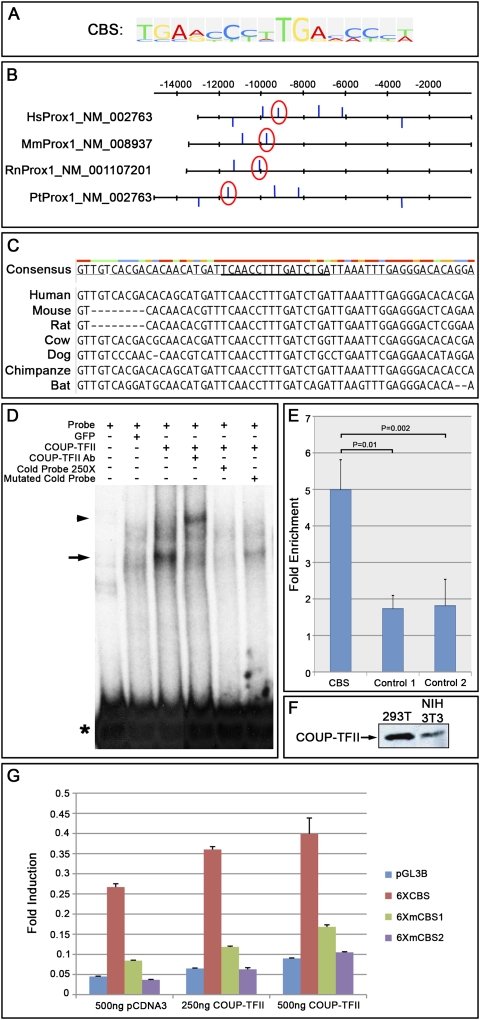
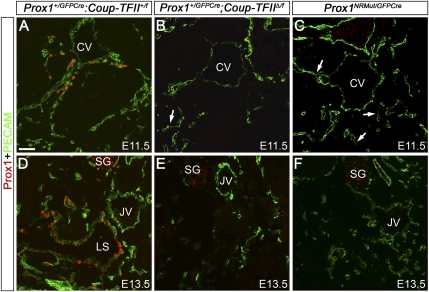
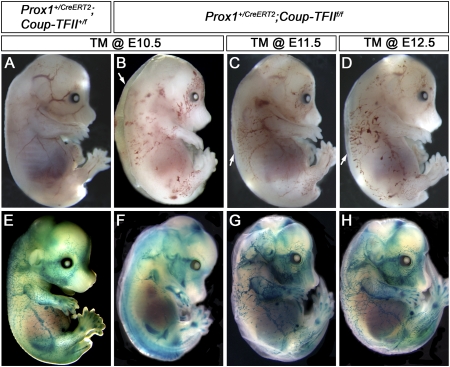
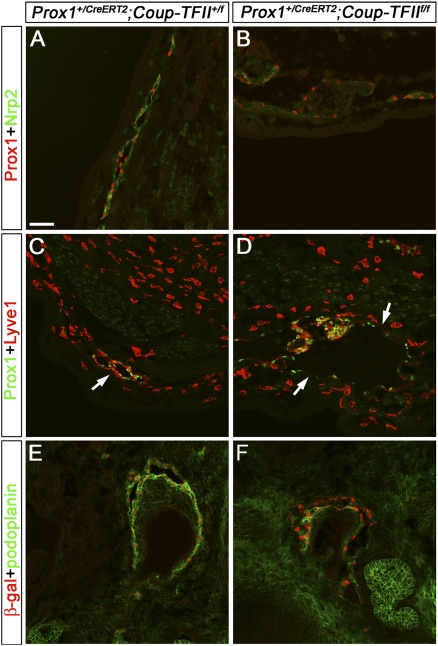
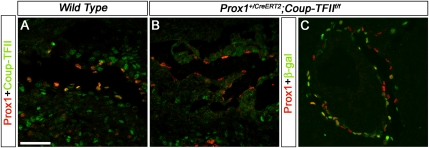
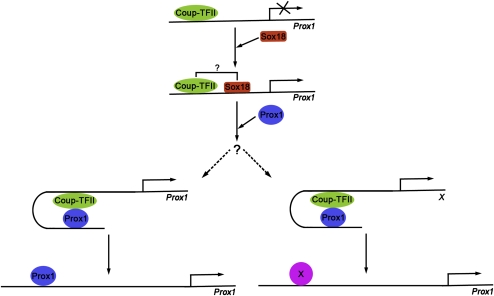
The homeobox gene Prox1 is crucial for mammalian lymphatic vascular development. In the absence of Prox1, lymphatic endothelial cells (LECs) are not specified. The maintenance of LEC identity also requires the constant expression of Prox1. However, the mechanisms controlling the expression of this gene in LECs remain poorly understood. The SRY-related gene Sox18 is required to induce Prox1 expression in venous LEC progenitors. Although Sox18 is also expressed in embryonic arteries, these vessels do not express Prox1, nor do they give rise to LECs. This finding suggests that some venous endothelial cell-specific factor is required for the activation of Prox1. Here we demonstrate that the nuclear hormone receptor Coup-TFII is necessary for the activation of Prox1 in embryonic veins by directly binding a conserved DNA domain in the regulatory region of Prox1. In addition, we show that the direct interaction between nuclear hormone receptors and Prox1 is also necessary for the maintenance of Prox1 expression during early stages of LEC specification and differentiation.
Figures
References
-
- de Boer J, Williams A, Skavdis G, Harker N, Coles M, Tolaini M, Norton T, Williams K, Roderick K, Potocnik AJ, et al. Transgenic mice with hematopoietic and lymphoid specific expression of Cre. Eur J Immunol. 2003;33:314–325. - PubMed
-
- Delorme B, Dahl E, Jarry-Guichard T, Briand JP, Willecke K, Gros D, Theveniau-Ruissy M. Expression pattern of connexin gene products at the early developmental stages of the mouse cardiovascular system. Circ Res. 1997;81:423–437. - PubMed
-
- Francois M, Caprini A, Hosking B, Orsenigo F, Wilhelm D, Browne C, Paavonen K, Karnezis T, Shayan R, Downes M, et al. Sox18 induces development of the lymphatic vasculature in mice. Nature. 2008;456:643–647. - PubMed
-
- Fruhwirth M, Janecke AR, Muller T, Carlton VE, Kronenberg F, Offner F, Knisely AS, Geleff S, Song EJ, Simma B, et al. Evidence for genetic heterogeneity in lymphedema-cholestasis syndrome. J Pediatr. 2003;142:441–447. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials