

T cell-dependence of Lassa fever pathogenesis
- PMID: 20360949
- PMCID: PMC2847900
- DOI: 10.1371/journal.ppat.1000836
T cell-dependence of Lassa fever pathogenesis
Abstract
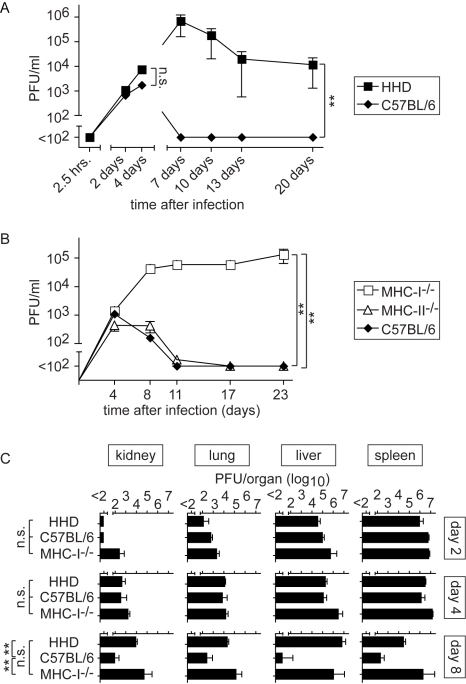
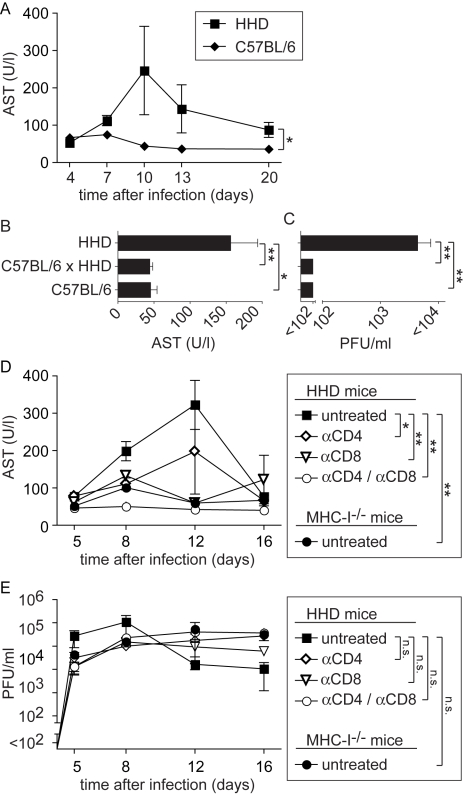
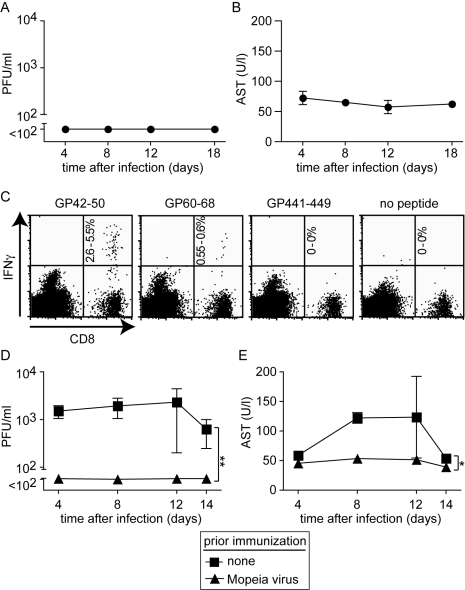
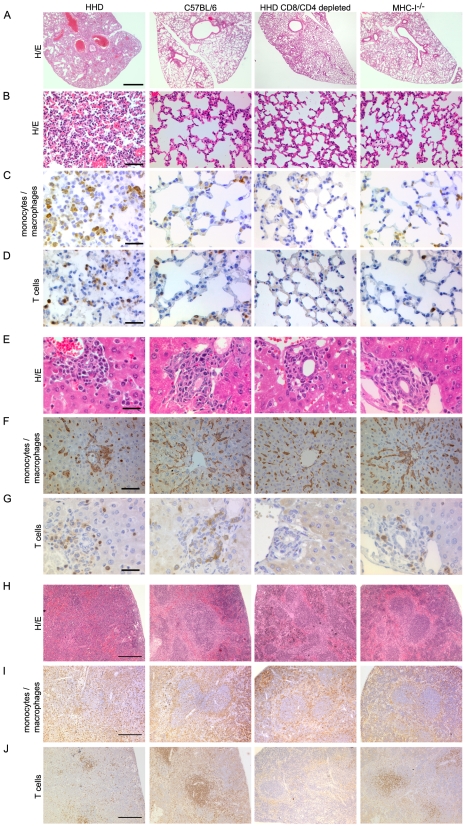
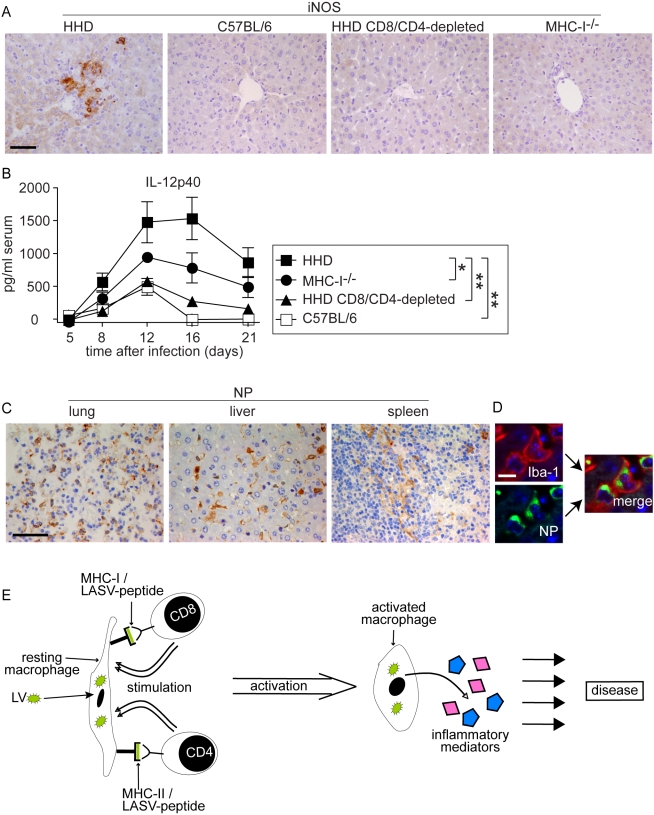
Lassa virus (LASV), the causative agent of Lassa fever (LF), is endemic in West Africa, accounting for substantial morbidity and mortality. In spite of ongoing research efforts, LF pathogenesis and mechanisms of LASV immune control remain poorly understood. While normal laboratory mice are resistant to LASV, we report that mice expressing humanized instead of murine MHC class I (MHC-I) failed to control LASV infection and develop severe LF. Infection of MHC-I knockout mice confirmed a key role for MHC-I-restricted T cell responses in controlling LASV. Intriguingly we found that T cell depletion in LASV-infected HHD mice prevented disease, irrespective of high-level viremia. Widespread activation of monocyte/macrophage lineage cells, manifest through inducible NO synthase expression, and elevated IL-12p40 serum levels indicated a systemic inflammatory condition. The absence of extensive monocyte/macrophage activation in T cell-depleted mice suggested that T cell responses contribute to deleterious innate inflammatory reactions and LF pathogenesis. Our observations in mice indicate a dual role for T cells, not only protecting from LASV, but also enhancing LF pathogenesis. The possibility of T cell-driven enhancement and immunopathogenesis should be given consideration in future LF vaccine development.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Buckley SM, Casals J, Downs WG. Isolation and antigenic characterization of Lassa virus. Nature. 1970;227:174. - PubMed
-
- McCormick JB, Webb PA, Krebs JW, Johnson KM, Smith ES. A prospective study of the epidemiology and ecology of Lassa fever. J Infect Dis. 1987;155:437–444. - PubMed
-
- Haas WH, Breuer T, Pfaff G, Schmitz H, Kohler P, et al. Imported Lassa fever in Germany: surveillance and management of contact persons. Clin Infect Dis. 2003;36:1254–1258. - PubMed
-
- Borio L, Inglesby T, Peters CJ, Schmaljohn AL, Hughes JM, et al. Hemorrhagic fever viruses as biological weapons: medical and public health management. Jama. 2002;287:2391–2405. - PubMed
-
- Charrel RN, de Lamballerie X. Arenaviruses other than Lassa virus. Antiviral Res. 2003;57:89–100. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
