

Origin and evolution of sulfadoxine resistant Plasmodium falciparum
- PMID: 20360965
- PMCID: PMC2847944
- DOI: 10.1371/journal.ppat.1000830
Origin and evolution of sulfadoxine resistant Plasmodium falciparum
Abstract
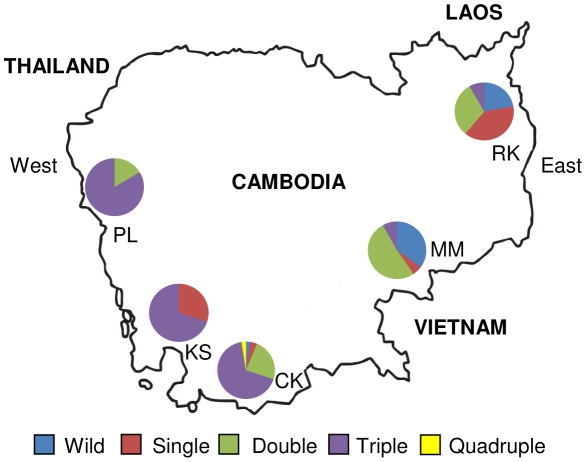
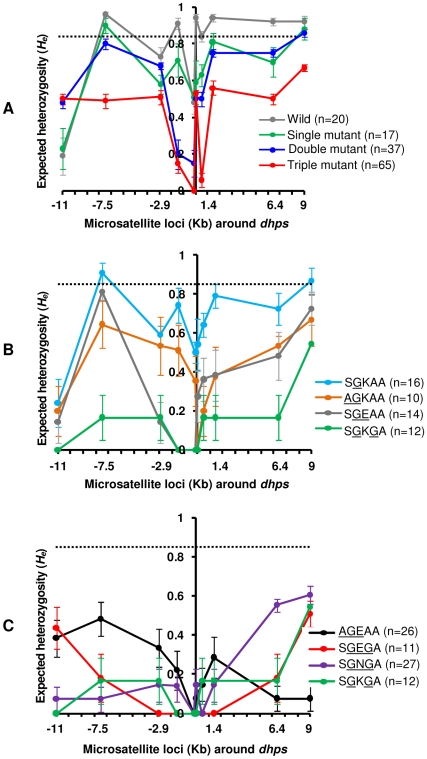
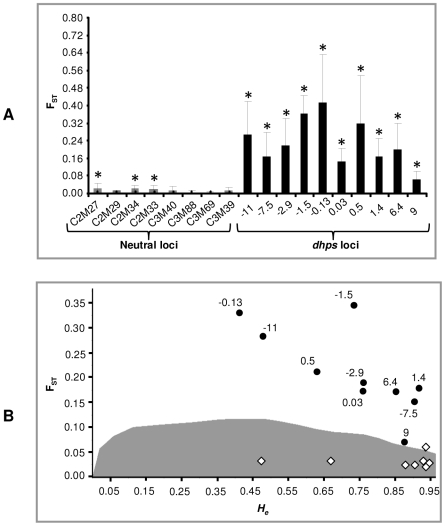
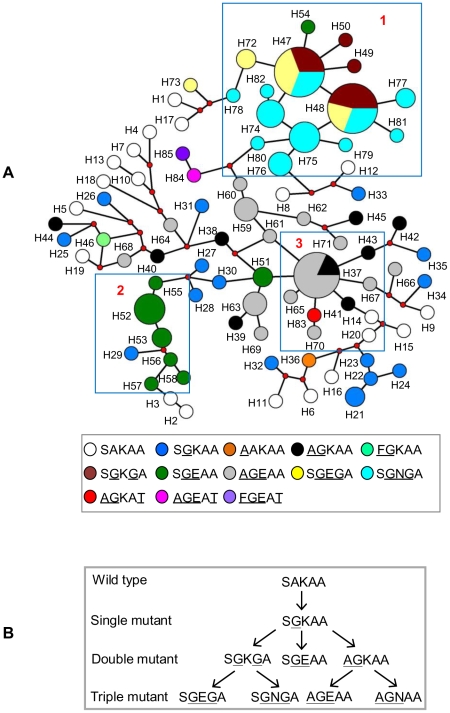
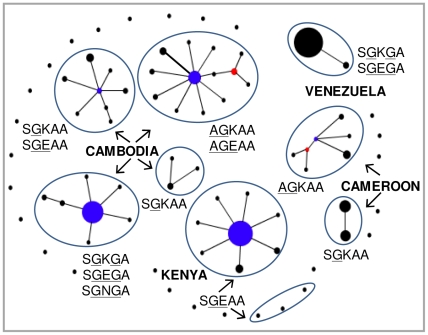
The Thailand-Cambodia border is the epicenter for drug-resistant falciparum malaria. Previous studies have shown that chloroquine (CQ) and pyrimethamine resistance originated in this region and eventually spread to other Asian countries and Africa. However, there is a dearth in understanding the origin and evolution of dhps alleles associated with sulfadoxine resistance. The present study was designed to reveal the origin(s) of sulfadoxine resistance in Cambodia and its evolutionary relationship to African and South American dhps alleles. We sequenced 234 Cambodian Plasmodium falciparum isolates for the dhps codons S436A/F, A437G, K540E, A581G and A613S/T implicated in sulfadoxine resistance. We also genotyped 10 microsatellite loci around dhps to determine the genetic backgrounds of various alleles and compared them with the backgrounds of alleles prevalent in Africa and South America. In addition to previously known highly-resistant triple mutant dhps alleles SGEGA and AGEAA (codons 436, 437, 540, 581, 613 are sequentially indicated), a large proportion of the isolates (19.3%) contained a 540N mutation in association with 437G/581G yielding a previously unreported triple mutant allele, SGNGA. Microsatellite data strongly suggest the strength of selection was greater on triple mutant dhps alleles followed by the double and single mutants. We provide evidence for at least three independent origins for the double mutants, one each for the SGKGA, AGKAA and SGEAA alleles. Our data suggest that the triple mutant allele SGEGA and the novel allele SGNGA have common origin on the SGKGA background, whereas the AGEAA triple mutant was derived from AGKAA on multiple, albeit limited, genetic backgrounds. The SGEAA did not share haplotypes with any of the triple mutants. Comparative analysis of the microsatellite haplotypes flanking dhps alleles from Cambodia, Kenya, Cameroon and Venezuela revealed an independent origin of sulfadoxine resistant alleles in each of these regions.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Eyles DE, Hoo CC, Warren M, Sandosham AA. Plasmodium falciparum resistant to chloroquine in Cambodia. Am J Trop Med Hyg. 1963;12:840–843. - PubMed
-
- Verdrager J. Epidemiology of the emergence and spread of drug-resistant falciparum malaria in South-East Asia and Australasia. J Trop Med Hyg. 1986;89:277–289. - PubMed
-
- Hurwitz ES, Johnson D, Campbell CC. Resistance of Plasmodium falciparum malaria to sulfadoxine-pyrimethamine (‘Fansidar’) in a refugee camp in Thailand. Lancet. 1981;1:1068–1070. - PubMed
-
- WHO. Development of South-Asia surveillance network for malaria drug resistance. 2002. Report on informal consultative meeting. World Health Organization, Geneva, Switzerland.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
