

The C-terminal tail of Yersinia pseudotuberculosis YopM is critical for interacting with RSK1 and for virulence
- PMID: 20368345
- PMCID: PMC2876544
- DOI: 10.1128/IAI.00141-10
The C-terminal tail of Yersinia pseudotuberculosis YopM is critical for interacting with RSK1 and for virulence
Abstract
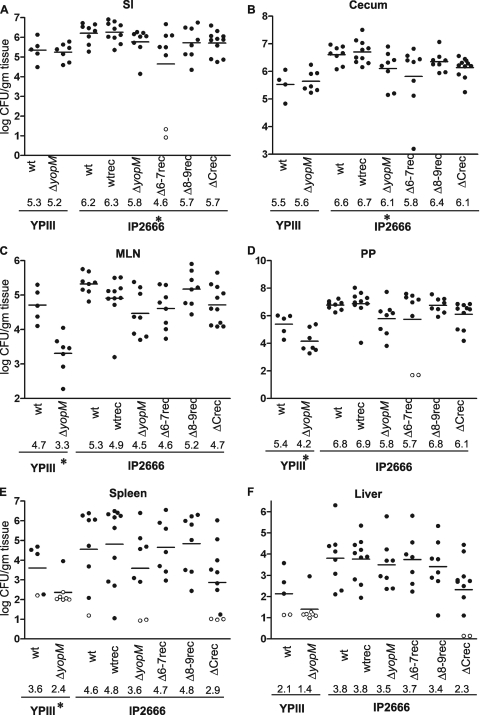
Yersinia spp. undermine the immune responses of infected animals by translocating Yops directly into host cells with a type III secretion system. YopM, a leucine-rich repeat protein, is a critical virulence factor in infection. YopM localizes to both the nucleus and the cytoplasm in cultured cells, interacts with mammalian p90 ribosomal S6 kinase 1 (RSK1), and causes a decrease in NK cell populations in spleens. Little is known about the molecular interaction between YopM and RSK1 and its significance in pathogenesis. We performed a systematic deletion analysis of YopM in Yersinia pseudotuberculosis to determine which regions are required for RSK1 interactions, nuclear localization, virulence, and changes in immune cell populations during infection of mice. Full-length YopM associated with RSK1 in at least two protein complexes in infected cells, and deletion of its C-terminal tail abrogated all RSK1 interactions. The C-terminal tail was required for tissue colonization, as yopM mutants that failed to interact with RSK1 were as defective for tissue colonization as was a DeltayopM mutant; however, nuclear localization of YopM was not dependent on its RSK1 interaction. Mutants expressing YopM proteins which do not interact with RSK1 caused more pathology than did the DeltayopM mutant, suggesting that there are other RSK1-independent functions of YopM. Histopathological and flow cytometric analyses of spleens showed that infection with wild-type Y. pseudotuberculosis caused an influx of neutrophils, while mice infected with yopM mutants had increased numbers of macrophages. Decreases in NK cells after Y. pseudotuberculosis infection did not correlate with YopM expression. In conclusion, the C terminus of YopM is essential for RSK1 interactions and for virulence.
Figures






 ), necrosis (
), necrosis ( ), and congestion (
), and congestion ( ). Scale bars represent 100 μm.
). Scale bars represent 100 μm.

References
-
- Arase, H., T. Saito, J. H. Phillips, and L. L. Lanier. 2001. Cutting edge: the mouse NK cell-associated antigen recognized by DX5 monoclonal antibody is CD49b (alpha 2 integrin, very late antigen-2). J. Immunol. 167:1141-1144. - PubMed
-
- Benabdillah, R., L. J. Mota, S. Lutzelschwab, E. Demoinet, and G. R. Cornelis. 2004. Identification of a nuclear targeting signal in YopM from Yersinia spp. Microb. Pathog. 36:247-261. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
