

Embryonic trafficking of gammadelta T cells to skin is dependent on E/P selectin ligands and CCR4
- PMID: 20368416
- PMCID: PMC2867765
- DOI: 10.1073/pnas.0912943107
Embryonic trafficking of gammadelta T cells to skin is dependent on E/P selectin ligands and CCR4
Abstract
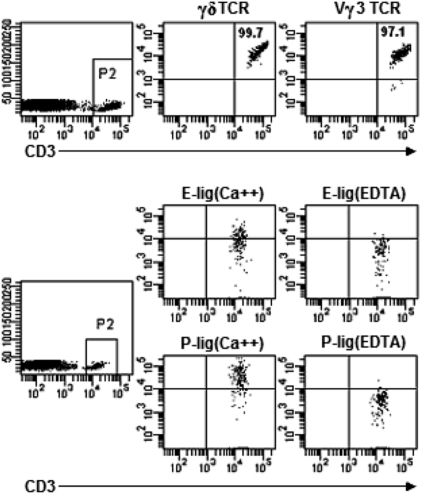
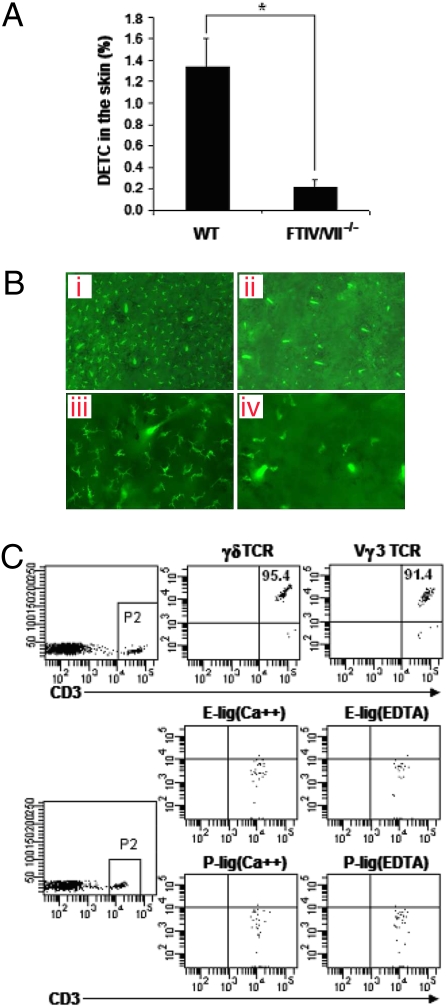
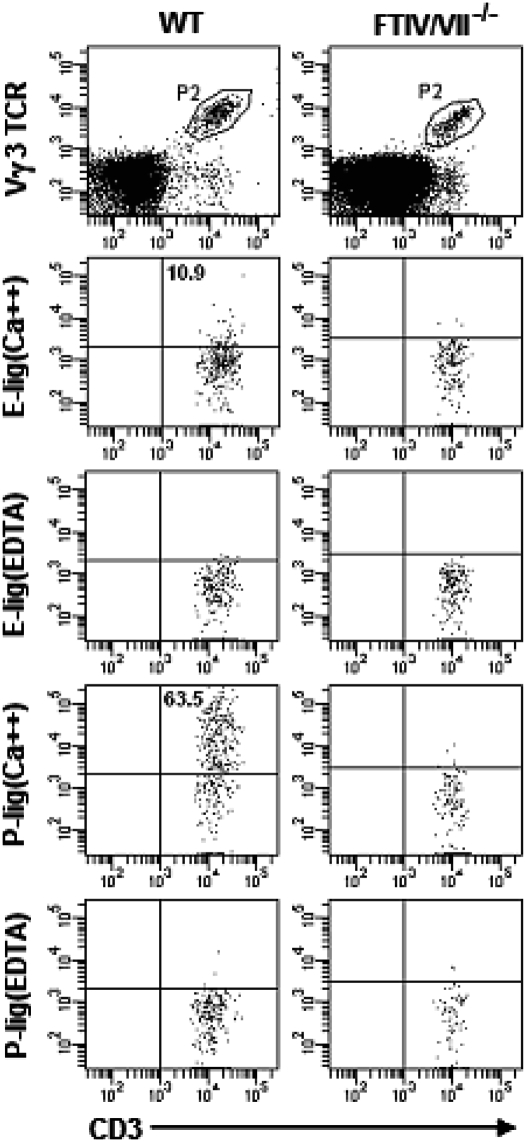
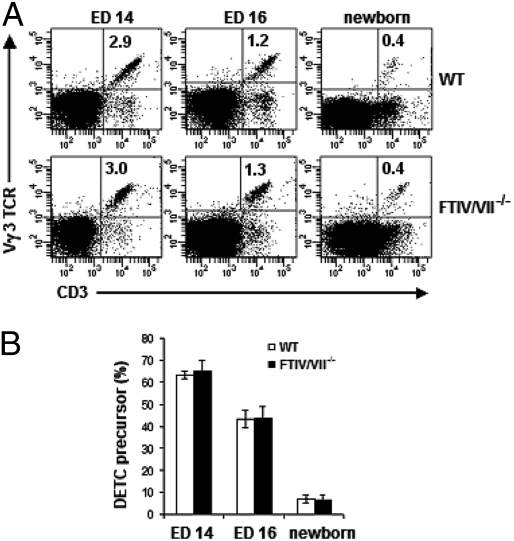
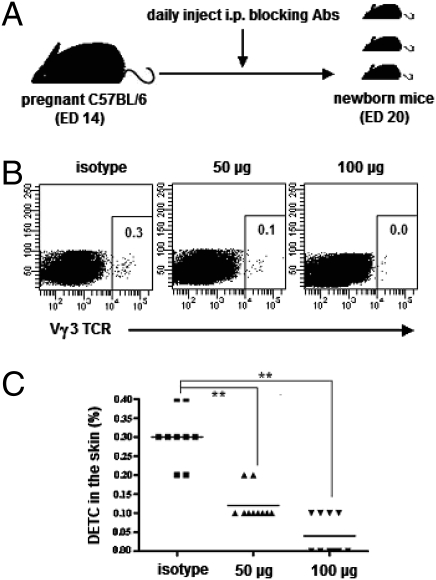
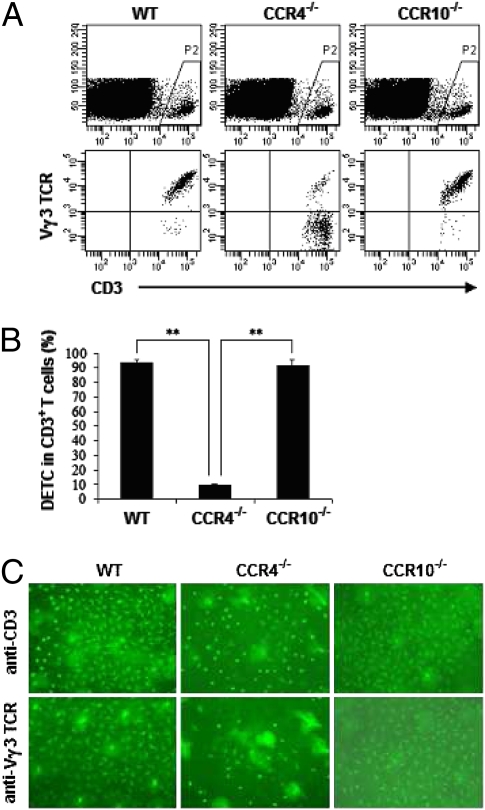
Dendritic epidermal T cells (DETC) express an invariant Vgamma3/Vdelta1 T-cell receptor, appear in fetal epidermis, and form a population of resident epidermal T cells. Their temporal development in the thymus has been studied extensively. However, little is known about the mechanisms involved in the embryonic trafficking of DETC from thymus to epidermis. We demonstrate that DETC in adult skin, as well as the DETC precursors in fetal thymus, express E and P selectin ligands (E- and P-lig). Mice deficient in alpha1,3 fucosyltransferases IV and VII (FTIV/VII) cannot synthesize the carbohydrate motifs that form key elements of these selectin ligands. The numbers of DETC in the epidermis of FTIV/VII(-/-) mice were dramatically reduced compared with normal mice. However, the development of DETC precursors in fetal thymus was identical in normal and FTIV/VII(-/-) mice, suggesting that the DETC precursors produced in FTIV/VII(-/-) mice could not traffic effectively to skin because they lack E- and P-lig. We tested this hypothesis by daily injection of blocking antibodies against E and P selectin into pregnant mice. Mice born from dams treated with anti-selectin antibodies, but not those born from dams treated with isotype control, had significantly diminished numbers of DETC. To test the role of chemokine receptors in DETC skin homing, we examined skin from CCR4(-/-) and CCR10(-/-) mice, respectively. DETC were significantly reduced in CCR4(-/-) mice but were present at normal levels in CCR10(-/-) mice. Our results present evidence for the crucial role of trafficking molecules in embryonic migration of DETC precursors to skin.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Saito H, et al. Complete primary structure of a heterodimeric T-cell receptor deduced from cDNA sequences. Nature. 1984;309:757–762. - PubMed
-
- Hayday AC, et al. Structure, organization, and somatic rearrangement of T cell gamma genes. Cell. 1985;40:259–269. - PubMed
-
- Brenner MB, et al. Identification of a putative second T-cell receptor. Nature. 1986;322:145–149. - PubMed
-
- Nanno M, Shiohara T, Yamamoto H, Kawakami K, Ishikawa H. gammadelta T cells: Firefighters or fire boosters in the front lines of inflammatory responses. Immunol Rev. 2007;215:103–113. - PubMed
-
- Payer E, Elbe A, Stingl G. Epidermal T lymphocytes—ontogeny, features and function. Springer Semin Immunopathol. 1992;13:315–331. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
