

Reaction mechanism of superoxide generation during ubiquinol oxidation by the cytochrome bc1 complex
- PMID: 20371599
- PMCID: PMC2878068
- DOI: 10.1074/jbc.M110.104364
Reaction mechanism of superoxide generation during ubiquinol oxidation by the cytochrome bc1 complex
Abstract
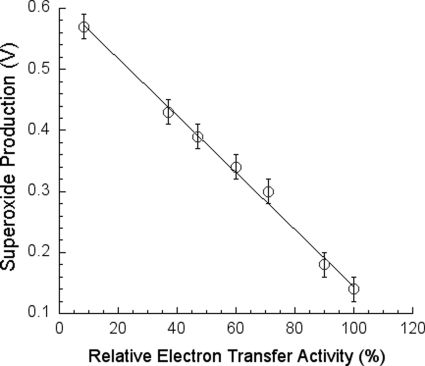
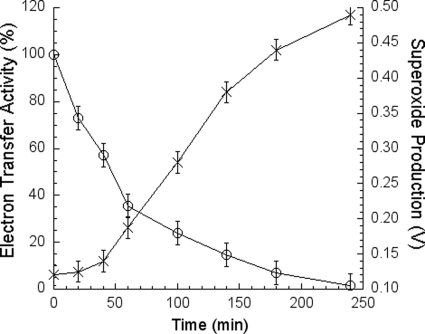
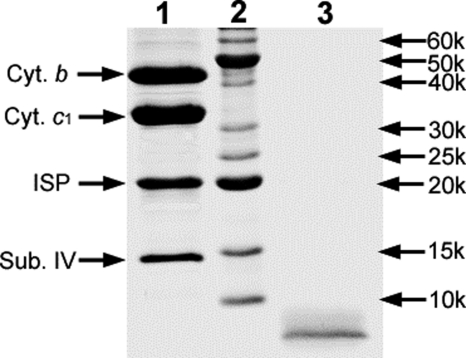
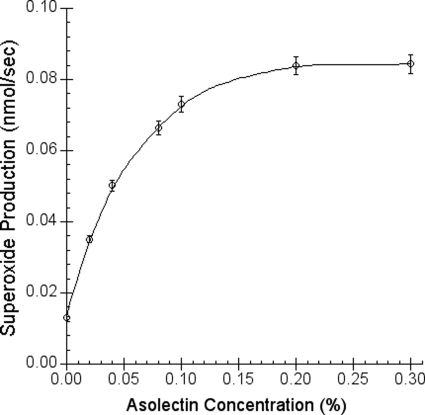
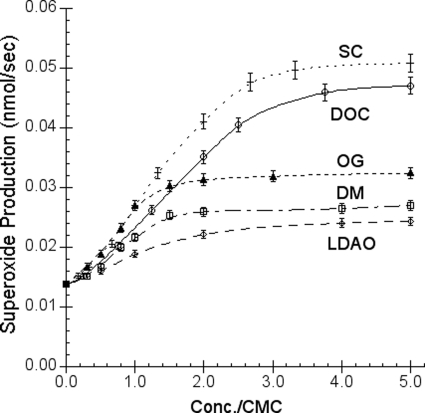
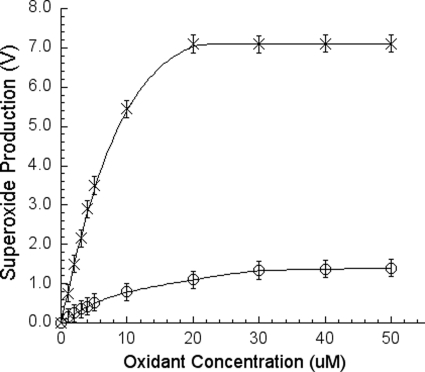
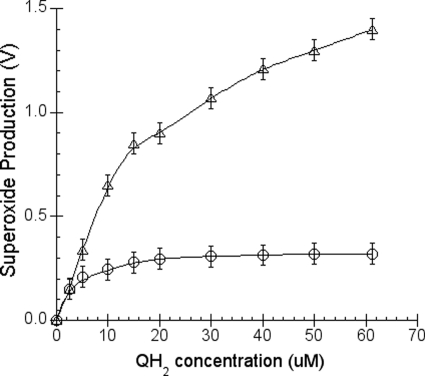
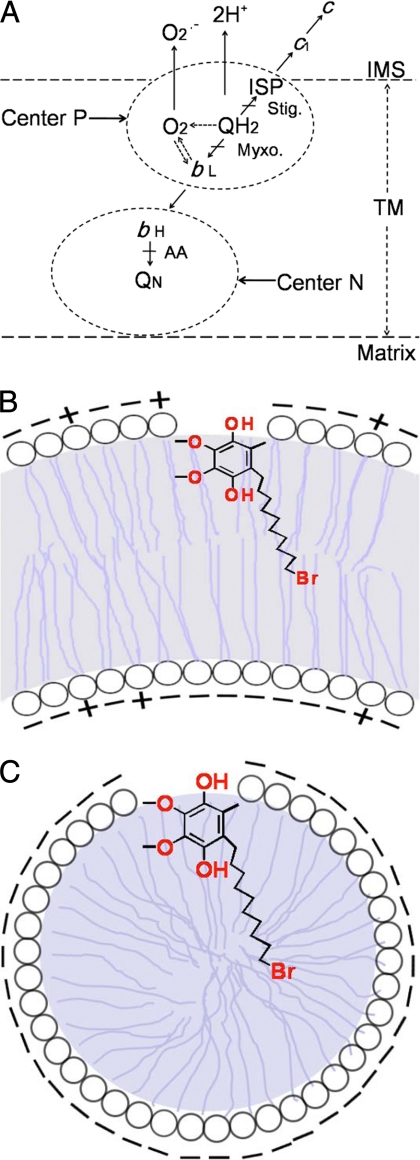
In addition to its main functions of electron transfer and proton translocation, the cytochrome bc(1) complex (bc(1)) also catalyzes superoxide anion (O(2)(*)) generation upon oxidation of ubiquinol in the presence of molecular oxygen. The reaction mechanism of superoxide generation by bc(1) remains elusive. The maximum O(2)(*) generation activity is observed when the complex is inhibited by antimycin A or inactivated by heat treatment or proteinase K digestion. The fact that the cytochrome bc(1) complex with less structural integrity has higher O(2)(*)-generating activity encouraged us to speculate that O(2)(*) is generated inside the complex, perhaps in the hydrophobic environment of the Q(P) pocket through bifurcated oxidation of ubiquinol by transferring its two electrons to a high potential electron acceptor, iron-sulfur cluster, and a low potential heme b(L) or molecular oxygen. If this speculation is correct, then one should see more O(2)(*) generation upon oxidation of ubiquinol by a high potential oxidant, such as cytochrome c or ferricyanide, in the presence of phospholipid vesicles or detergent micelles than in the hydrophilic conditions, and this is indeed the case. The protein subunits, at least those surrounding the Q(P) pocket, may play a role either in preventing the release of O(2)(*) from its production site to aqueous environments or in preventing O(2) from getting access to the hydrophobic Q(P) pocket and might not directly participate in superoxide production.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
