

Identification and functional characterization of a novel mitochondrial carrier for citrate and oxoglutarate in Saccharomyces cerevisiae
- PMID: 20371607
- PMCID: PMC2878499
- DOI: 10.1074/jbc.M109.097188
Identification and functional characterization of a novel mitochondrial carrier for citrate and oxoglutarate in Saccharomyces cerevisiae
Abstract
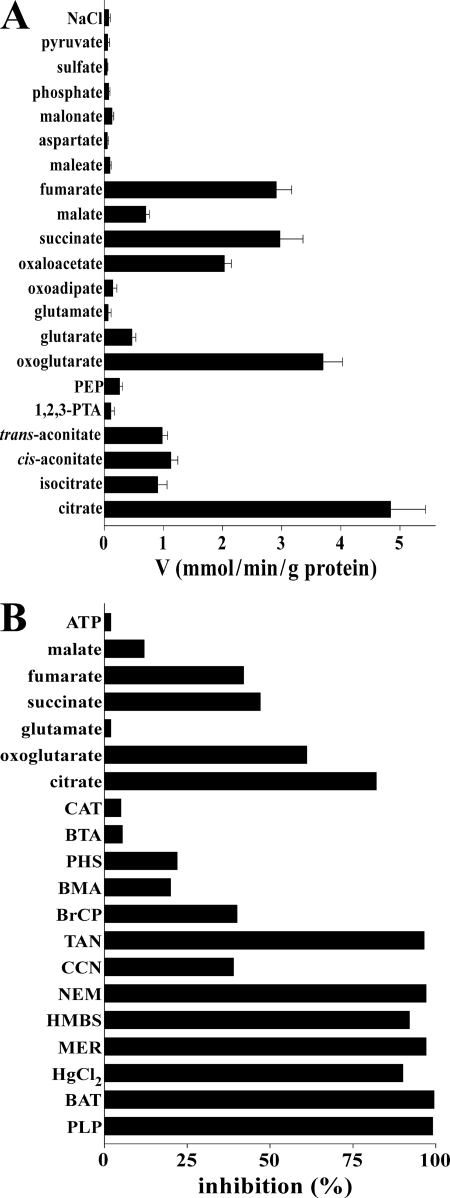
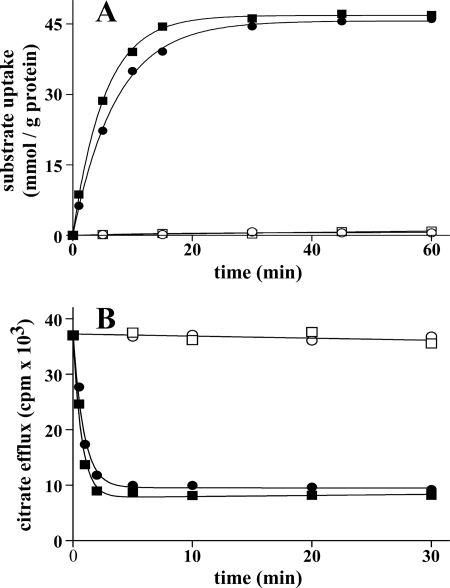
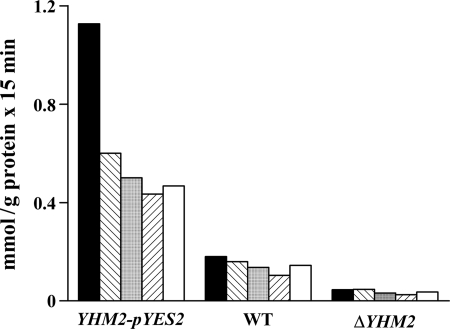
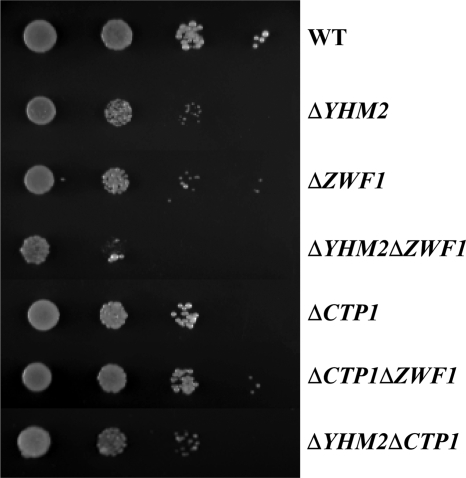
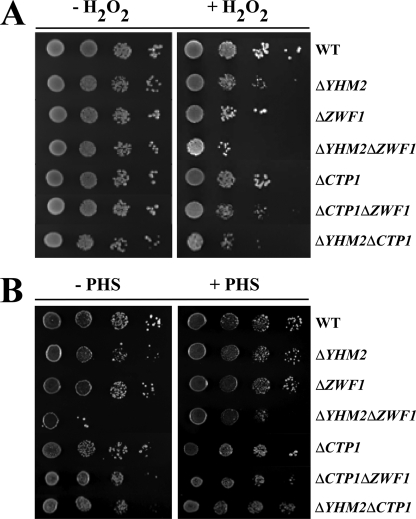
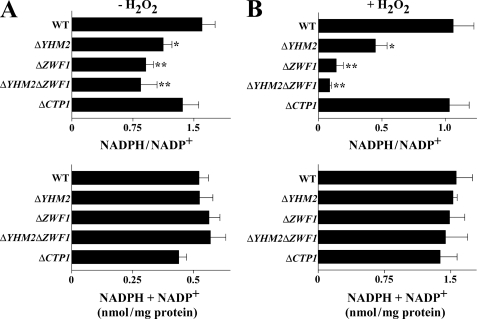
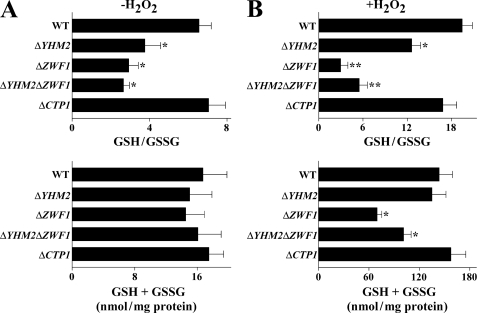
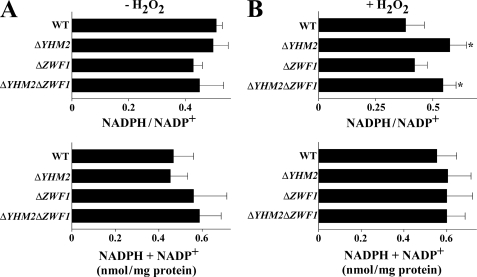
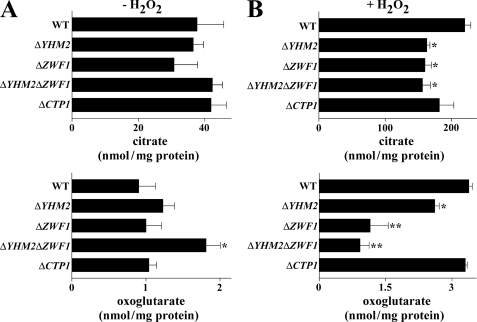
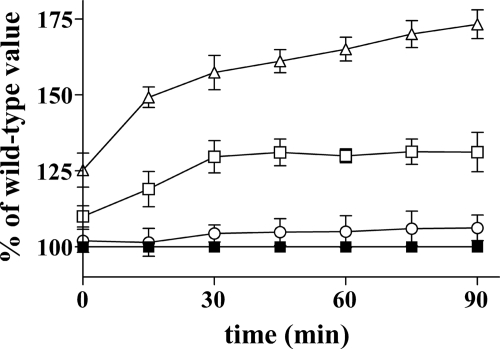
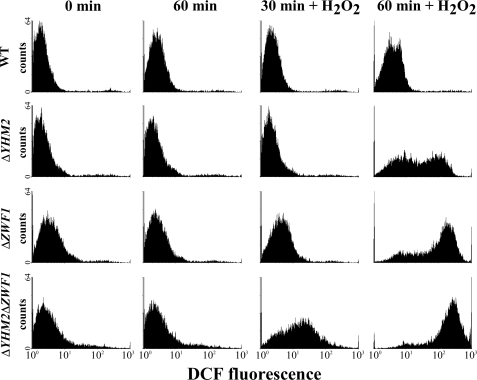
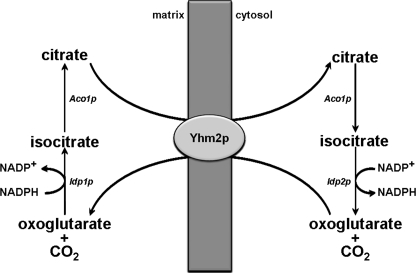
Mitochondrial carriers are a family of transport proteins that shuttle metabolites, nucleotides, and coenzymes across the mitochondrial membrane. The function of only a few of the 35 Saccharomyces cerevisiae mitochondrial carriers still remains to be uncovered. In this study, we have functionally defined and characterized the S. cerevisiae mitochondrial carrier Yhm2p. The YHM2 gene was overexpressed in S. cerevisiae, and its product was purified and reconstituted into liposomes. Its transport properties, kinetic parameters, and targeting to mitochondria show that Yhm2p is a mitochondrial transporter for citrate and oxoglutarate. Reconstituted Yhm2p also transported oxaloacetate, succinate, and fumarate to a lesser extent, but virtually not malate and isocitrate. Yhm2p catalyzed only a counter-exchange transport that was saturable and inhibited by sulfhydryl-blocking reagents but not by 1,2,3-benzenetricarboxylate (a powerful inhibitor of the citrate/malate carrier). The physiological role of Yhm2p is to increase the NADPH reducing power in the cytosol (required for biosynthetic and antioxidant reactions) and probably to act as a key component of the citrate-oxoglutarate NADPH redox shuttle between mitochondria and cytosol. This protein function is based on observations documenting a decrease in the NADPH/NADP(+) and GSH/GSSG ratios in the cytosol of DeltaYHM2 cells as well as an increase in the NADPH/NADP(+) ratio in their mitochondria compared with wild-type cells. Our proposal is also supported by the growth defect displayed by the DeltaYHM2 strain and more so by the DeltaYHM2DeltaZWF1 strain upon H(2)O(2) exposure, implying that Yhm2p has an antioxidant function.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
