

Beyond 9+0: noncanonical axoneme structures characterize sensory cilia from protists to humans
- PMID: 20371625
- PMCID: PMC2923350
- DOI: 10.1096/fj.09-151381
Beyond 9+0: noncanonical axoneme structures characterize sensory cilia from protists to humans
Abstract
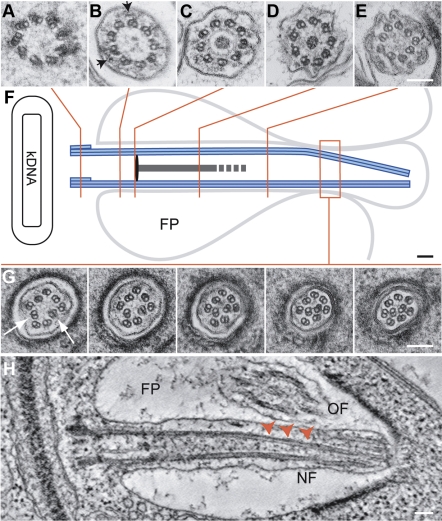
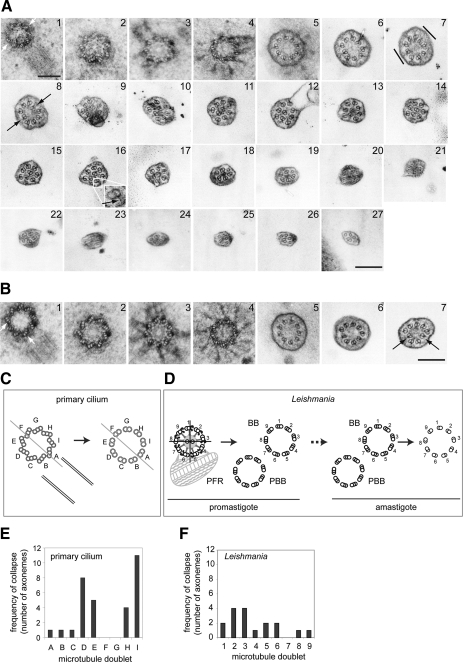
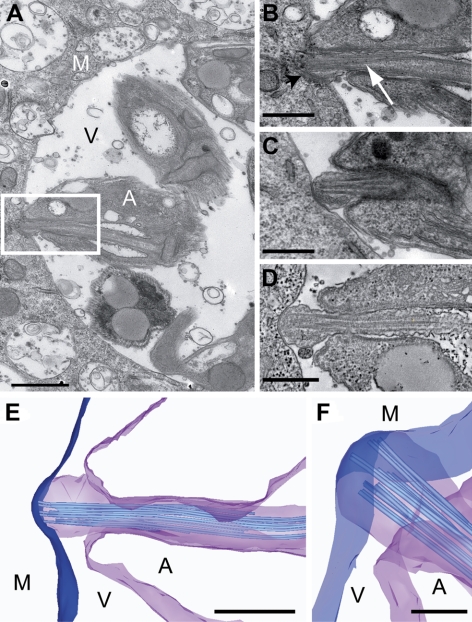
The intracellular amastigote stages of parasites such as Leishmania are often referred to as aflagellate. They do, however, possess a short axoneme of cryptic function. Here, our examination of the structure of this axoneme leads to a testable hypothesis of its role in the cell biology of pathogenicity. We show a striking similarity between the microtubule axoneme structure of the Leishmania mexicana parasite infecting a macrophage and vertebrate primary cilia. In both, the 9-fold microtubule doublet symmetry is broken by the incursion of one or more microtubule doublets into the axoneme core, giving rise to an architecture that we term here the 9v (variable) axoneme. Three-dimensional reconstructions revealed that no particular doublet initiated the symmetry break, and moreover it often involved 2 doublets. The tip of the L. mexicana flagellum was frequently intimately associated with the macrophage vacuole membrane. We propose that the main function of the amastigote flagellum is to act as a sensory organelle with important functions in host-parasite interactions and signaling in the intracellular stage of the L. mexicana life cycle.
Figures
References
-
- Gull K. The parasite point of view: insights and questions on the cell biology of Trypanosoma and Leishmania parasite-phagocyte interactions. Russell D G, Gordon S, editors. Washington, DC: ASM Press; Phagocyte-Pathogen Interactions: Macrophages and the Host Response to Infection. 2009:453–462.
-
- Simpson C F, Harvey J W, French T W. Ultrastructure of amastigotes of Leishmania donovani in the bone marrow of a dog. Am J Vet Res. 1982;43:1684–1686. - PubMed
-
- Alexander J. Unusual axonemal doublet arrangements in the flagellum of Leishmania amastigotes. Trans R Soc Trop Med Hyg. 1978;72:345–347. - PubMed
-
- Hentzer B, Kobayasi T. The ultrastructure of Leishmania tropica in skin lesions. Acta Pathol Microbiol Scand B. 1977;85:153–160. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
