

Nuclear transport and the mitotic apparatus: an evolving relationship
- PMID: 20372967
- PMCID: PMC11115906
- DOI: 10.1007/s00018-010-0325-7
Nuclear transport and the mitotic apparatus: an evolving relationship
Abstract
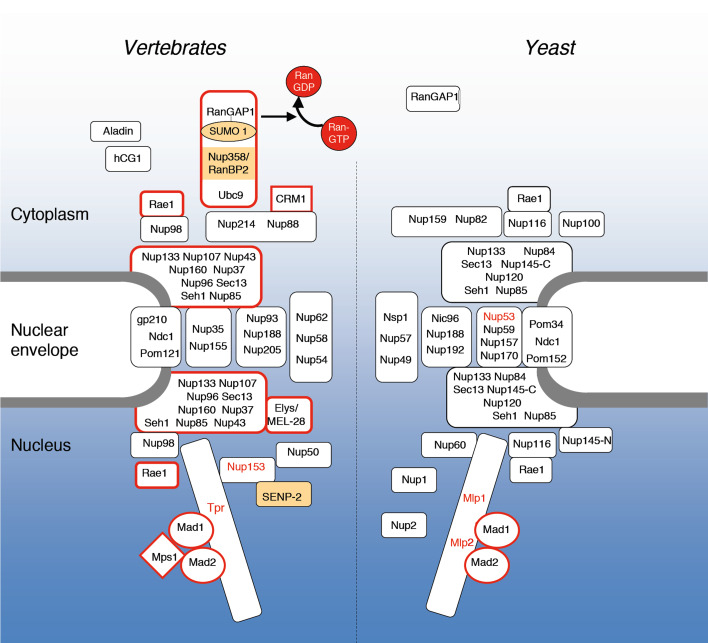
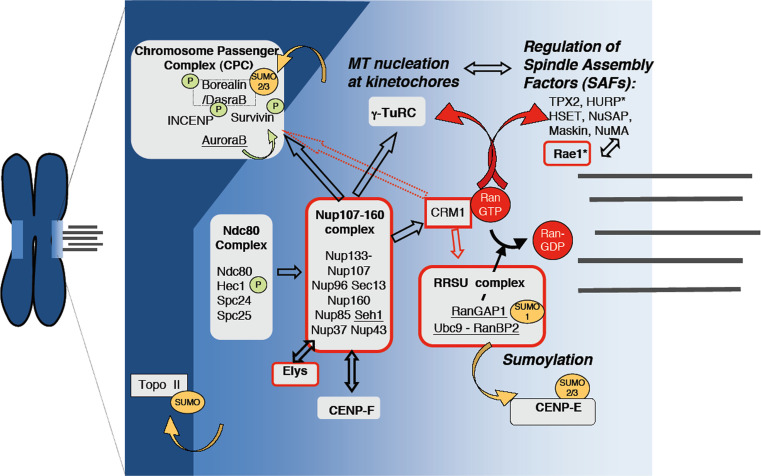
The trafficking of macromolecules between the cytoplasm and the nucleus is controlled by the nuclear pore complexes (NPCs) and various transport factors that facilitate the movement of cargos through the NPCs and their accumulation in the target compartment. While their functions in transport are well established, an ever-growing number of observations have also linked components of the nuclear transport machinery to processes that control chromosome segregation during mitosis, including spindle assembly, kinetochore function, and the spindle assembly checkpoint. In this review, we will discuss this evolving area of study and emerging hypotheses that propose key roles for components of the nuclear transport apparatus in mitotic progression.
Figures

References
-
- Tran EJ, Wente SR. Dynamic nuclear pore complexes: life on the edge. Cell. 2006;125:1041–1053. - PubMed
-
- Antonin W, Ellenberg J, Dultz E. Nuclear pore complex assembly through the cell cycle: regulation and membrane organization. FEBS Lett. 2008;582:2004–2016. - PubMed
-
- Chan GK, Liu ST, Yen TJ. Kinetochore structure and function. Trends Cell Biol. 2005;15:589–598. - PubMed
-
- Maiato H, DeLuca J, Salmon ED, Earnshaw WC. The dynamic kinetochore-microtubule interface. J Cell Sci. 2004;117:5461–5477. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
