

The effects of hemodynamic force on embryonic development
- PMID: 20374481
- PMCID: PMC2927969
- DOI: 10.1111/j.1549-8719.2010.00025.x
The effects of hemodynamic force on embryonic development
Abstract
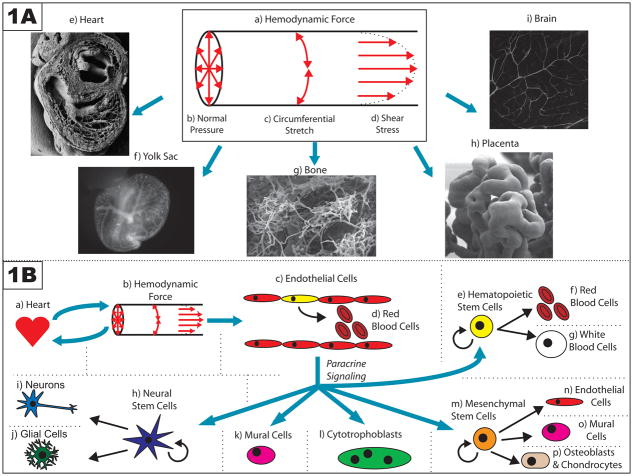
Blood vessels have long been known to respond to hemodynamic force, and several mechanotransduction pathways have been identified. However, only recently have we begun to understand the effects of hemodynamic force on embryonic development. In this review, we will discuss specific examples illustrating the role of hemodynamic force during the development of the embryo, with particular focus on the development of the vascular system and the morphogenesis of the heart. We will also discuss the important functions served by mechanotransduction and hemodynamic force during placentation, as well as in regulating the maintenance and division of embryonic, hematopoietic, neural, and mesenchymal stem cells. Pathological misregulation of mechanosensitive pathways during pregnancy and embryonic development may contribute to the occurrence of cardiovascular birth defects, as well as to a variety of other diseases, including preeclampsia. Thus, there is a need for future studies focusing on better understanding the physiological effects of hemodynamic force during embryonic development and their role in the pathogenesis of disease.
Figures


References
-
- Aikawa R, Nagai T, Kudoh S, Zou Y, Tanaka M, Tamura M, Akazawa H, Takano H, Nagai R, Komuro I. Integrins play a critical role in mechanical stress-induced p38 MAPK activation. Hypertension. 2002;39:233–238. - PubMed
-
- Armulik A, Abramsson A, Betsholtz C. Endothelial/pericyte interactions. Circ Res. 2005;97:512–523. - PubMed
-
- Arvidsson A, Collin T, Kirik D, Kokaia Z, Lindvall O. Neuronal replacement from endogenous precursors in the adult brain after stroke. Nat Med. 2002;8:963–970. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
