

Euryarchaeal beta-CASP proteins with homology to bacterial RNase J Have 5'- to 3'-exoribonuclease activity
- PMID: 20375016
- PMCID: PMC2878522
- DOI: 10.1074/jbc.M109.095117
Euryarchaeal beta-CASP proteins with homology to bacterial RNase J Have 5'- to 3'-exoribonuclease activity
Abstract
In the Archaea only a handful of ribonucleases involved in RNA processing and degradation have been characterized. One potential group of archaeal ribonucleases are homologues of the bacterial RNase J family, which have a beta-CASP metallo-beta-lactamase fold. Here we show that beta-CASP proteins encoded in the genomes of the hyperthermophilic Euryarchaeota Pyrococcus abyssi and Thermococcus kodakaraensis are processive exoribonucleases with a 5' end dependence and a 5' to 3' directionality. We named these enzymes Pab-RNase J and Tk-RNase J, respectively. RNAs with 5'-monophosphate or 5'-hydroxyl ends are preferred substrates of Pab-RNase J, whereas circularized RNA is resistant to Pab-RNase J activity. Degradation of a 3' end-labeled synthetic RNA in which an internal nucleoside is substituted by three ethylene glycol units generates intermediates demonstrating 5' to 3' directionality. The substitution of conserved residues in Pab-RNase J predicted to be involved in the coordination of metal ions demonstrates their importance for ribonuclease activity, although the detailed geometry of the catalytic site is likely to differ from bacterial RNase J. This is the first identification of a 5'-exoribonuclease encoded in the genomes of the Archaea. Phylogenetic analysis shows that euryarchaeal RNase J has been inherited vertically, suggesting an ancient origin predating the separation of the Bacteria and the Archaea.
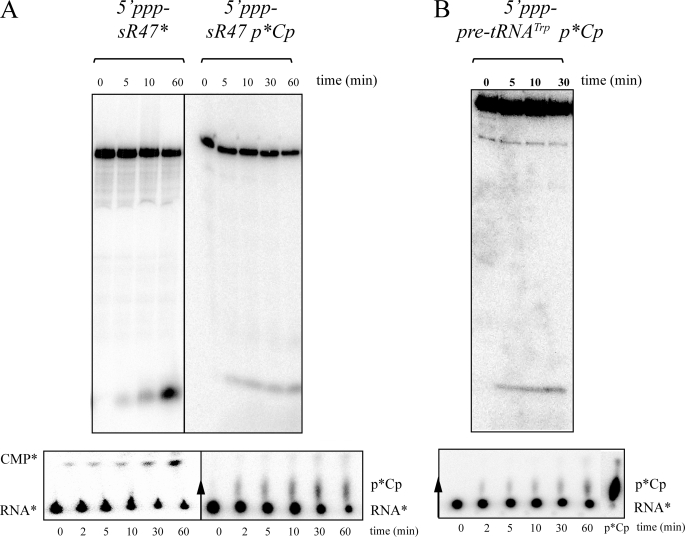
Figures






References
-
- Evguenieva-Hackenberg E., Klug G. (2009) Prog. Mol. Biol. Transl. Sci. 85, 275–317 - PubMed
-
- Frank D. N., Pace N. R. (1998) Annu. Rev. Biochem. 67, 153–180 - PubMed
-
- Redko Y., Li de Lasierra-Gallay I., Condon C. (2007) Nat. Rev. Microbiol. 5, 278–286 - PubMed
-
- Li H., Trotta C. R., Abelson J. (1998) Science 280, 279–284 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
