

Spatial representation of alarm pheromone information in a secondary olfactory centre in the ant brain
- PMID: 20375054
- PMCID: PMC2894929
- DOI: 10.1098/rspb.2010.0366
Spatial representation of alarm pheromone information in a secondary olfactory centre in the ant brain
Abstract
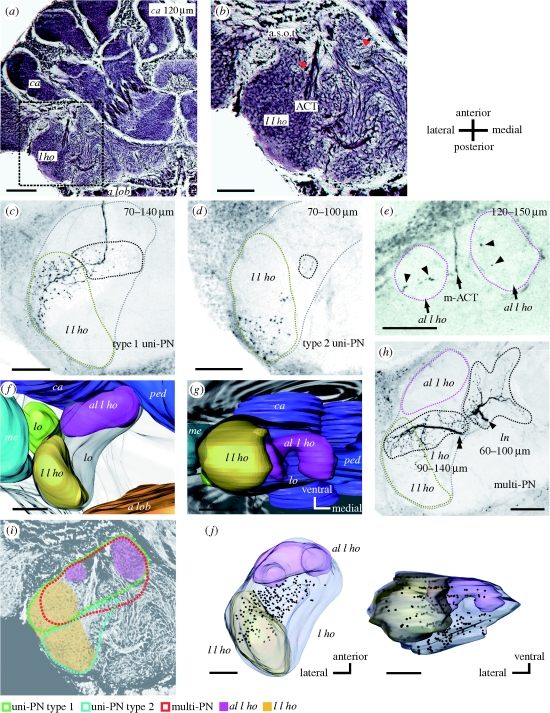
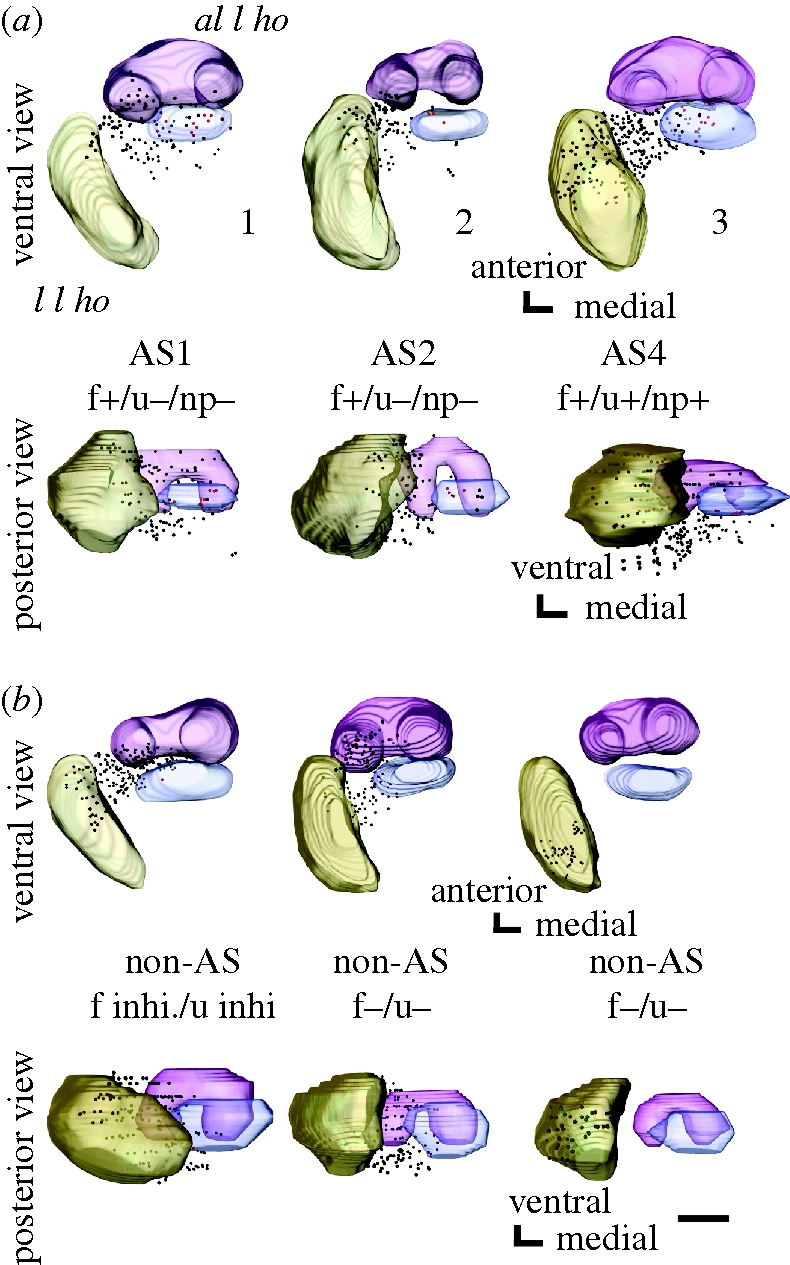
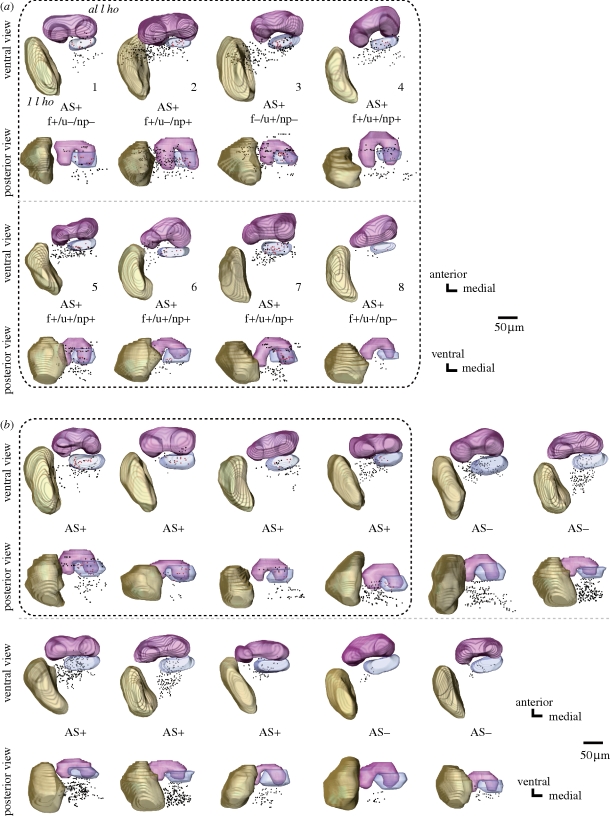
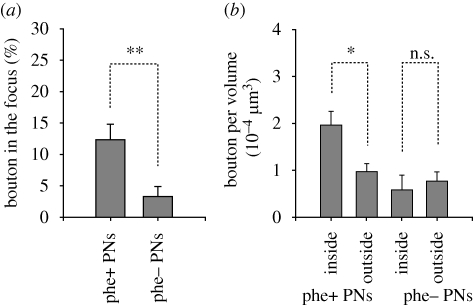
Pheromones play major roles in intraspecific communication in many animals. Elaborated communication systems in eusocial insects provide excellent materials to study neural mechanisms for social pheromone processing. We previously reported that alarm pheromone information is processed in a specific cluster of glomeruli in the antennal lobe of the ant Camponotus obscuripes. However, representation of alarm pheromone information in a secondary olfactory centre is unknown in any animal. Olfactory information in the antennal lobe is transmitted to secondary olfactory centres, including the lateral horn, by projection neurons (PNs). In this study, we compared distributions of terminal boutons of alarm pheromone-sensitive and -insensitive PNs in the lateral horn of ants. Distributions of their dendrites largely overlapped, but there was a region where boutons of pheromone-sensitive PNs, but not those of pheromone-insensitive PNs, were significantly denser than in the rest of the lateral horn. Moreover, most of a major type of pheromone-sensitive efferent neurons from the lateral horn extended dendritic branches in this region, suggesting specialization of this region for alarm pheromone processing. This study is the first study to demonstrate the presence of specialized areas for the processing of a non-sexual, social pheromone in the secondary olfactory centre in any animal.
Figures
Similar articles
-
Alarm pheromone processing in the ant brain: an evolutionary perspective.Front Behav Neurosci. 2010 Jun 8;4:28. doi: 10.3389/fnbeh.2010.00028. eCollection 2010. Front Behav Neurosci. 2010. PMID: 20676235 Free PMC article.
-
Pheromone-sensitive glomeruli in the primary olfactory centre of ants.Proc Biol Sci. 2006 Sep 7;273(1598):2219-25. doi: 10.1098/rspb.2006.3565. Proc Biol Sci. 2006. PMID: 16901842 Free PMC article.
-
Neural pathways for the processing of alarm pheromone in the ant brain.J Comp Neurol. 2007 Dec 1;505(4):424-42. doi: 10.1002/cne.21500. J Comp Neurol. 2007. PMID: 17912739
-
Neural mechanisms of alarm pheromone signaling.Mol Cells. 2013 Mar;35(3):177-81. doi: 10.1007/s10059-013-0056-3. Epub 2013 Mar 5. Mol Cells. 2013. PMID: 23471444 Free PMC article. Review.
-
Central mechanisms of pheromone information processing.Chem Senses. 1996 Apr;21(2):269-75. doi: 10.1093/chemse/21.2.269. Chem Senses. 1996. PMID: 8670705 Review.
Cited by
-
Alarm pheromone processing in the ant brain: an evolutionary perspective.Front Behav Neurosci. 2010 Jun 8;4:28. doi: 10.3389/fnbeh.2010.00028. eCollection 2010. Front Behav Neurosci. 2010. PMID: 20676235 Free PMC article.
-
Behavioral and neurophysiological study of olfactory perception and learning in honeybees.Front Syst Neurosci. 2011 Dec 8;5:98. doi: 10.3389/fnsys.2011.00098. eCollection 2011. Front Syst Neurosci. 2011. PMID: 22163215 Free PMC article.
-
Representation of pheromones, interspecific signals, and plant odors in higher olfactory centers; mapping physiologically identified antennal-lobe projection neurons in the male heliothine moth.Front Syst Neurosci. 2014 Oct 9;8:186. doi: 10.3389/fnsys.2014.00186. eCollection 2014. Front Syst Neurosci. 2014. PMID: 25346663 Free PMC article.
-
Functional analysis of a higher olfactory center, the lateral horn.J Neurosci. 2012 Jun 13;32(24):8138-48. doi: 10.1523/JNEUROSCI.1066-12.2012. J Neurosci. 2012. PMID: 22699895 Free PMC article.
References
-
- Boeckh J., Ernst K. D.1987Contribution of single unit analysis in insects to an understanding of olfactory function. J. Comp. Physiol. A. 161, 549–565 (doi:10.1007/BF00603661) - DOI
-
- Brennan P. A., Kendrick K. M.2006Mammalian social odours: attraction and individual recognition. Phil. Trans. R. Soc. B. 361, 2061–2078 (doi:10.1098/rstb.2006.1931) - DOI - PMC - PubMed
-
- Brennan P. A., Zufall F.2006Pheromonal communication in vertebrates. Nature 444, 308–315 (doi:10.1038/nature05404) - DOI - PubMed
-
- Couto A., Alenius M., Dickson B. J.2005Molecular, anatomical, and functional organization of the Drosophila olfactory system. Curr. Biol. 15, 1535–1547 (doi:10.1016/j.cub.2005.07.034) - DOI - PubMed
-
- de Belle J. S., Heisenberg M.1994Associative odor learning in Drosophila abolished by chemical ablation of mushroom bodies. Science 263, 692–695 (doi:10.1126/science.8303280) - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
