

Character displacement from the receiver's perspective: species and mate recognition despite convergent signals in suboscine birds
- PMID: 20375056
- PMCID: PMC2894922
- DOI: 10.1098/rspb.2010.0210
Character displacement from the receiver's perspective: species and mate recognition despite convergent signals in suboscine birds
Abstract
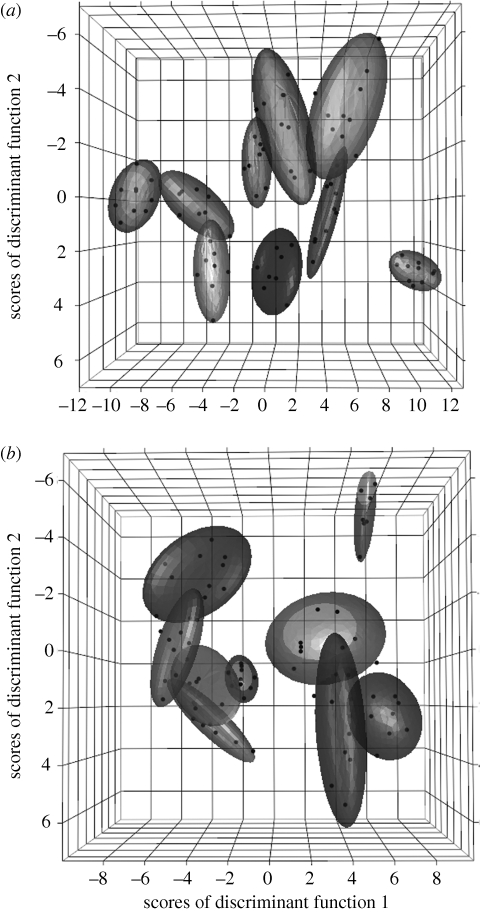
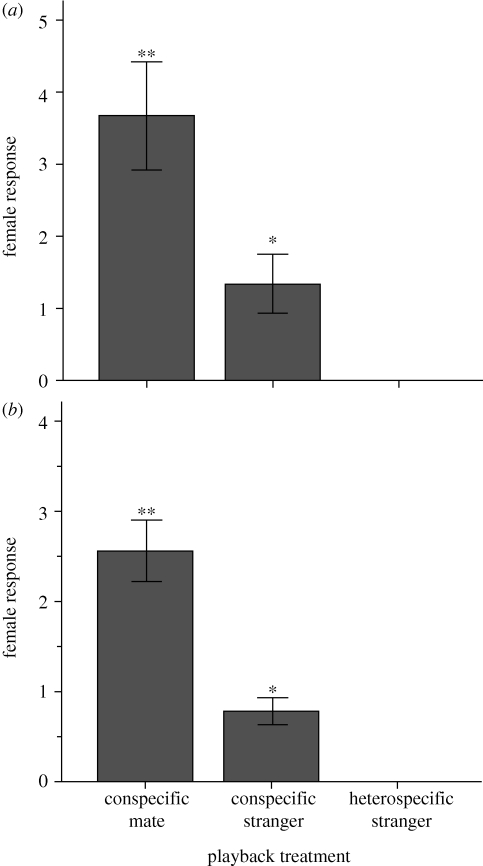
Many social animals use long-distance signals to attract mates and defend territories. They face the twin challenges of discriminating between species to identify conspecific mates, and between individuals to recognize collaborators and competitors. It is therefore often assumed that long-distance signals are under strong selection for species-specificity and individual distinctiveness, and that this will drive character displacement when closely related species meet, particularly in noisy environments. However, the occurrence of signal stereotypy and convergence in rainforest species seems to contradict these ideas, and raises the question of whether receivers in these systems can recognize species or individuals by long-distance signals alone. Here, we test for acoustically mediated recognition in two sympatric antbird species that are known to have convergent songs. We show that male songs are stereotyped yet individually distinctive, and we use playback experiments to demonstrate that females can discriminate not only between conspecific and heterospecific males, but between mates and strangers. These findings provide clear evidence that stereotypy and convergence in male signals can be accommodated by fine tuning of perceptual abilities in female receivers, suggesting that the evolutionary forces driving divergent character displacement in animal signals are weaker than is typically assumed.
Figures
Similar articles
-
Ecological adaptation and species recognition drives vocal evolution in neotropical suboscine birds.Evolution. 2005 Jan;59(1):200-15. Evolution. 2005. PMID: 15792239
-
Range-wide spatial mapping reveals convergent character displacement of bird song.Proc Biol Sci. 2019 May 15;286(1902):20190443. doi: 10.1098/rspb.2019.0443. Proc Biol Sci. 2019. PMID: 31039717 Free PMC article.
-
Deciphering information encoded in birdsong: male songbirds with fertile mates respond most strongly to complex, low-amplitude songs used in courtship.Am Nat. 2011 Oct;178(4):478-87. doi: 10.1086/661901. Epub 2011 Aug 15. Am Nat. 2011. PMID: 21956026
-
Neural mechanisms of auditory species recognition in birds.Biol Rev Camb Philos Soc. 2019 Oct;94(5):1619-1635. doi: 10.1111/brv.12518. Epub 2019 May 7. Biol Rev Camb Philos Soc. 2019. PMID: 31066222 Review.
-
To Become Senders, Songbirds Must be Receivers First.Integr Comp Biol. 2017 Oct 1;57(4):910-919. doi: 10.1093/icb/icx106. Integr Comp Biol. 2017. PMID: 28985314 Review.
Cited by
-
Reproductive character displacement: insights from genital morphometrics in damselfly hybrid zones.Heredity (Edinb). 2024 Nov;133(5):355-368. doi: 10.1038/s41437-024-00719-9. Epub 2024 Aug 18. Heredity (Edinb). 2024. PMID: 39155287
-
Captive rearing experiments confirm song development without learning in a tracheophone suboscine bird.PLoS One. 2014 Apr 30;9(4):e95746. doi: 10.1371/journal.pone.0095746. eCollection 2014. PLoS One. 2014. PMID: 24788343 Free PMC article.
-
Asymmetric Response of Costa Rican White-Breasted Wood-Wrens (Henicorhina leucosticta) to Vocalizations from Allopatric Populations.PLoS One. 2015 Dec 15;10(12):e0144949. doi: 10.1371/journal.pone.0144949. eCollection 2015. PLoS One. 2015. PMID: 26671001 Free PMC article.
-
Do zebra finch parents fail to recognise their own offspring?PLoS One. 2011 Apr 13;6(4):e18466. doi: 10.1371/journal.pone.0018466. PLoS One. 2011. PMID: 21533233 Free PMC article.
-
Interspecific competition, hybridization, and reproductive isolation in secondary contact: missing perspectives on males and females.Curr Zool. 2018 Feb;64(1):75-88. doi: 10.1093/cz/zox060. Epub 2017 Oct 28. Curr Zool. 2018. PMID: 29492041 Free PMC article.
References
-
- Baptista L. F., Kroodsma D. E.2001Avian bioacoustics. In Handbook of the birds of the world (eds del Hoyo J., Elliott A., Sargatal J.), pp. 11–52 Barcelona, Spain: Lynx Edicions
-
- Bard S. C., Hau M., Wikelski M., Wingfield J. C.2002Vocal distinctiveness and response to conspecific playback in the spotted antbird, a Neotropical suboscine. Condor 104, 387–394 (doi:10.1650/0010-5422(2002)104[0387:VDARTC]2.0.CO;2) - DOI
-
- Becker P. H.1982The coding of species-specific characteristics in bird sounds. In Acoustic communication in birds (eds Kroodsma D. E., Miller E. H.), pp. 213–252 New York, NY: Academic Press
-
- Bee M. A., Gerhardt H. C.2002Individual voice recognition in a territorial frog (Rana catesbeiana). Proc. R. Soc. B 269, 1443–1448 (doi:10.1098/rspb.2002.2041) - DOI - PMC - PubMed
-
- Beecher M. D., Brenowitz E. A.2005Functional aspects of song learning in songbirds. Trends Ecol. Evol. 20, 143–150 (doi:10.1016/j.tree.2005.01.004) - DOI - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
