

Virus cell-to-cell transmission
- PMID: 20375157
- PMCID: PMC2918988
- DOI: 10.1128/JVI.00443-10
Virus cell-to-cell transmission
Abstract
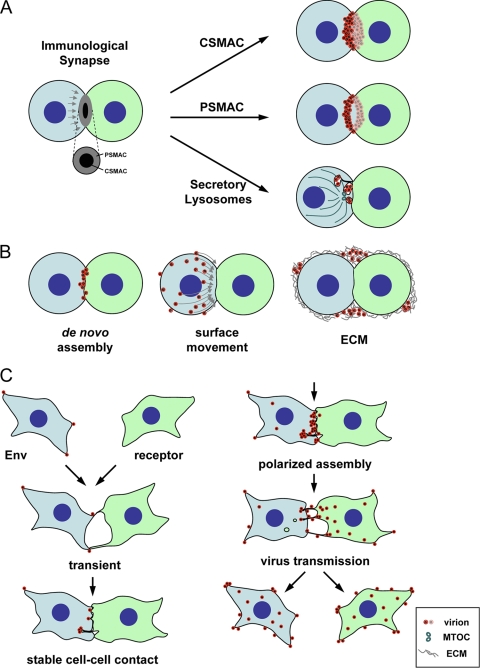
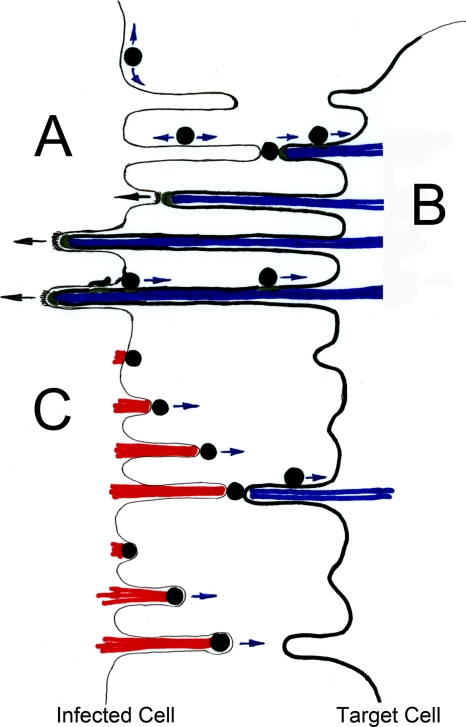
Viral infections spread based on the ability of viruses to overcome multiple barriers and move from cell to cell, tissue to tissue, and person to person and even across species. While there are fundamental differences between these types of transmissions, it has emerged that the ability of viruses to utilize and manipulate cell-cell contact contributes to the success of viral infections. Central to the excitement in the field of virus cell-to-cell transmission is the idea that cell-to-cell spread is more than the sum of the processes of virus release and entry. This implies that virus release and entry are efficiently coordinated to sites of cell-cell contact, resulting in a process that is distinct from its individual components. In this review, we will present support for this model, illustrate the ability of viruses to utilize and manipulate cell adhesion molecules, and discuss the mechanism and driving forces of directional spreading. An understanding of viral cell-to-cell spreading will enhance our ability to intervene in the efficient spreading of viral infections.
Figures
References
-
- Abe, K., O. Chisaka, F. Van Roy, and M. Takeichi. 2004. Stability of dendritic spines and synaptic contacts is controlled by alpha N-catenin. Nat. Neurosci. 7:357-363. - PubMed
-
- Agosto, L. M., J. J. Yu, M. K. Liszewski, C. Baytop, N. Korokhov, L. M. Humeau, and U. O'Doherty. 2009. The CXCR4-tropic human immunodeficiency virus envelope promotes more-efficient gene delivery to resting CD4+ T cells than the vesicular stomatitis virus glycoprotein G envelope. J. Virol. 83:8153-8162. - PMC - PubMed
-
- Akins, M. R., and T. Biederer. 2006. Cell-cell interactions in synaptogenesis. Curr. Opin. Neurobiol. 16:83-89. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
