

Effect of mutations in VP5 hydrophobic loops on rotavirus cell entry
- PMID: 20375171
- PMCID: PMC2876642
- DOI: 10.1128/JVI.02461-09
Effect of mutations in VP5 hydrophobic loops on rotavirus cell entry
Abstract
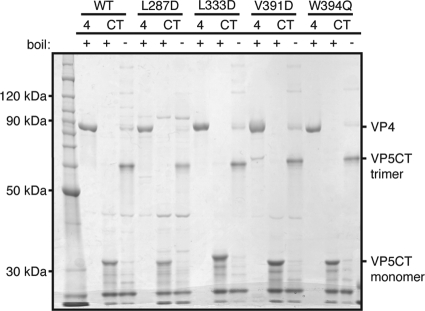
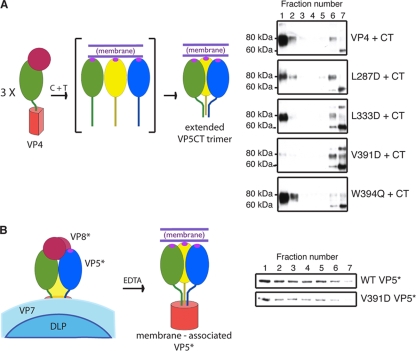
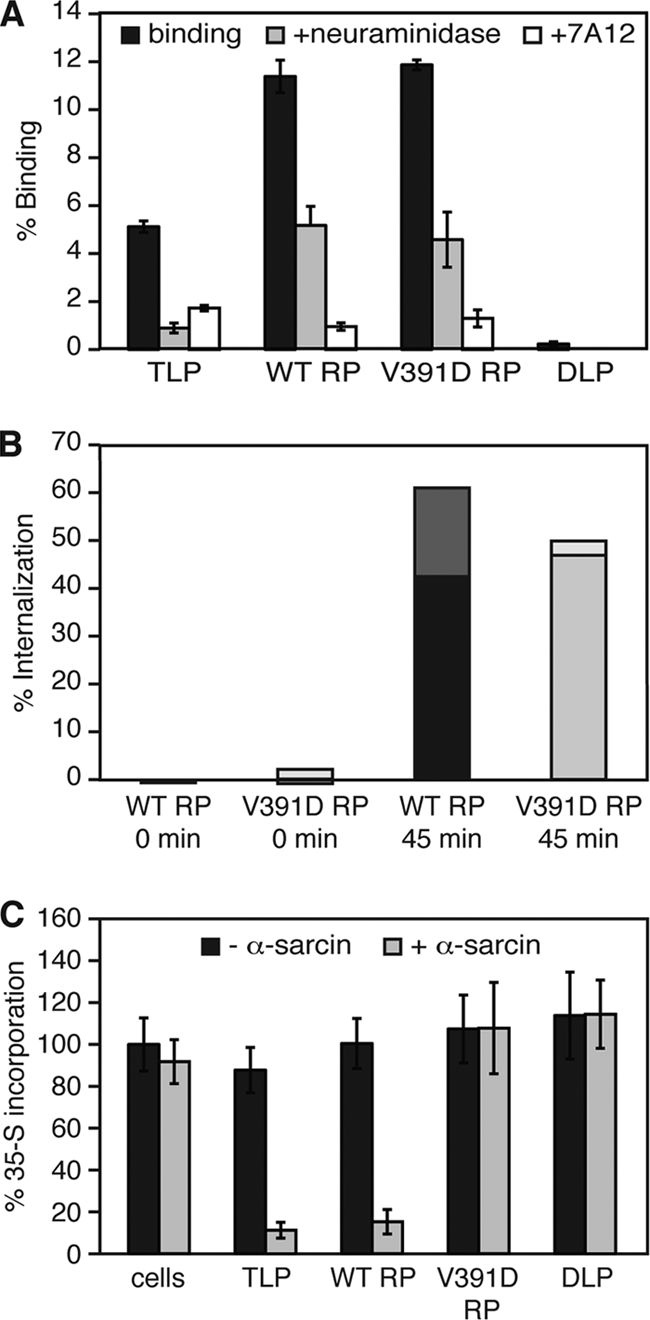
Experiments in cell-free systems have demonstrated that the VP5 cleavage fragment of the rotavirus spike protein, VP4, undergoes a foldback rearrangement that translocates three clustered hydrophobic loops from one end of the molecule to the other. This conformational change resembles the foldback rearrangements of enveloped virus fusion proteins. By recoating rotavirus subviral particles with recombinant VP4 and VP7, we tested the effects on cell entry of substituting hydrophilic for hydrophobic residues in the clustered VP5 loops. Several of these mutations decreased the infectivity of recoated particles without preventing either recoating or folding back. In particular, the V391D mutant had a diminished capacity to interact with liposomes when triggered to fold back by serial protease digestion in solution, and particles recoated with this mutant VP4 were 10,000-fold less infectious than particles recoated with wild-type VP4. Particles with V391D mutant VP4 attached normally to cells and internalized efficiently, but they failed in the permeabilization step that allows coentry of the toxin alpha-sarcin. These findings indicate that the hydrophobicity of the VP5 apex is required for membrane disruption during rotavirus cell entry.
Figures
Similar articles
-
A rotavirus spike protein conformational intermediate binds lipid bilayers.J Virol. 2010 Feb;84(4):1764-70. doi: 10.1128/JVI.01682-09. Epub 2009 Dec 9. J Virol. 2010. PMID: 20007281 Free PMC article.
-
VP5* rearranges when rotavirus uncoats.J Virol. 2009 Nov;83(21):11372-7. doi: 10.1128/JVI.01228-09. Epub 2009 Aug 19. J Virol. 2009. PMID: 19692464 Free PMC article.
-
Assembly of highly infectious rotavirus particles recoated with recombinant outer capsid proteins.J Virol. 2006 Nov;80(22):11293-304. doi: 10.1128/JVI.01346-06. Epub 2006 Sep 13. J Virol. 2006. PMID: 16971442 Free PMC article.
-
Rotavirus Replication: Gaps of Knowledge on Virus Entry and Morphogenesis.Tohoku J Exp Med. 2019 Aug;248(4):285-296. doi: 10.1620/tjem.248.285. Tohoku J Exp Med. 2019. PMID: 31447474 Review.
-
A hypothesis about the mechanism of assembly of double-shelled rotavirus particles.Arch Virol Suppl. 1996;12:79-85. doi: 10.1007/978-3-7091-6553-9_9. Arch Virol Suppl. 1996. PMID: 9015104 Review.
Cited by
-
Virus Type-Specific Removal in a Full-Scale Membrane Bioreactor Treatment Process.Food Environ Virol. 2018 Jun;10(2):176-186. doi: 10.1007/s12560-017-9330-4. Epub 2017 Dec 6. Food Environ Virol. 2018. PMID: 29214559
-
Functional refolding of the penetration protein on a non-enveloped virus.Nature. 2021 Feb;590(7847):666-670. doi: 10.1038/s41586-020-03124-4. Epub 2021 Jan 13. Nature. 2021. PMID: 33442061 Free PMC article.
-
Comparative analysis of Reoviridae reverse genetics methods.Methods. 2013 Feb;59(2):199-206. doi: 10.1016/j.ymeth.2012.05.012. Epub 2012 Jun 8. Methods. 2013. PMID: 22687622 Free PMC article. Review.
-
Mutations in the rotavirus spike protein VP4 reduce trypsin sensitivity but not viral spread.J Gen Virol. 2013 Jun;94(Pt 6):1296-1300. doi: 10.1099/vir.0.050674-0. Epub 2013 Feb 20. J Gen Virol. 2013. PMID: 23426355 Free PMC article.
-
Atomic model of an infectious rotavirus particle.EMBO J. 2011 Jan 19;30(2):408-16. doi: 10.1038/emboj.2010.322. Epub 2010 Dec 14. EMBO J. 2011. PMID: 21157433 Free PMC article.
References
-
- Bong, D. T., C. Steinem, A. Janshoff, J. E. Johnson, and M. Reza Ghadiri. 1999. A highly membrane-active peptide in Flock House virus: implications for the mechanism of nodavirus infection. Chem. Biol. 6:473-481. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
