

The evolutionary history of the Arabidopsis lyrata complex: a hybrid in the amphi-Beringian area closes a large distribution gap and builds up a genetic barrier
- PMID: 20377907
- PMCID: PMC2858744
- DOI: 10.1186/1471-2148-10-98
The evolutionary history of the Arabidopsis lyrata complex: a hybrid in the amphi-Beringian area closes a large distribution gap and builds up a genetic barrier
Abstract
Background: The genomes of higher plants are, on the majority, polyploid, and hybridisation is more frequent in plants than in animals. Both polyploidisation and hybridisation contribute to increased variability within species, and may transfer adaptations between species in a changing environment. Studying these aspects of evolution within a diversified species complex could help to clarify overall spatial and temporal patterns of plant speciation. The Arabidopsis lyrata complex, which is closely related to the model plant Arabidopsis thaliana, is a perennial, outcrossing, herbaceous species complex with a circumpolar distribution in the Northern Hemisphere as well as a disjunct Central European distribution in relictual habitats. This species complex comprises three species and four subspecies, mainly diploids but also several tetraploids, including one natural hybrid. The complex is ecologically, but not fully geographically, separated from members of the closely related species complex of Arabidopsis halleri, and the evolutionary histories of both species compexes have largely been influenced by Pleistocene climate oscillations.
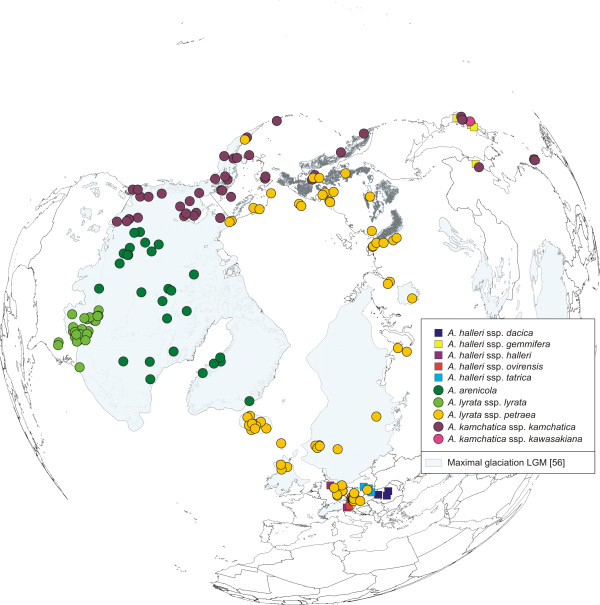
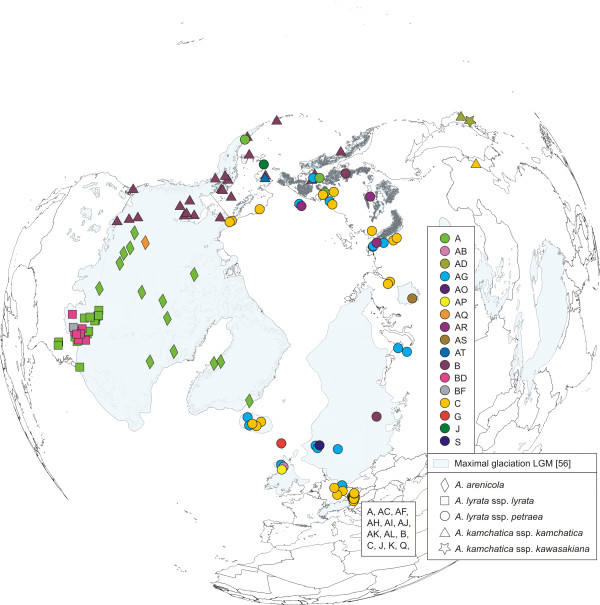
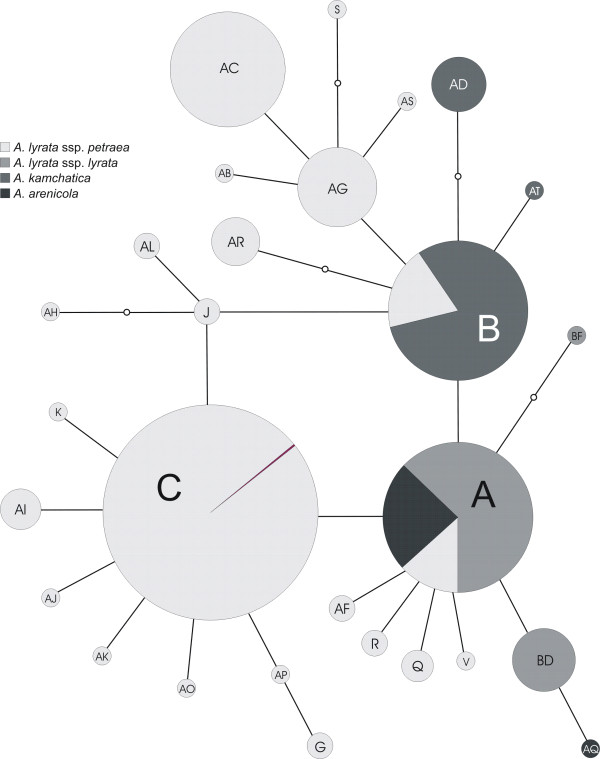
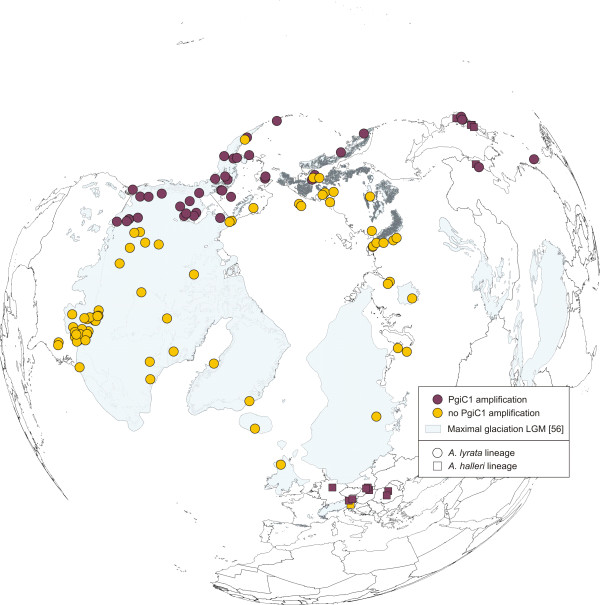
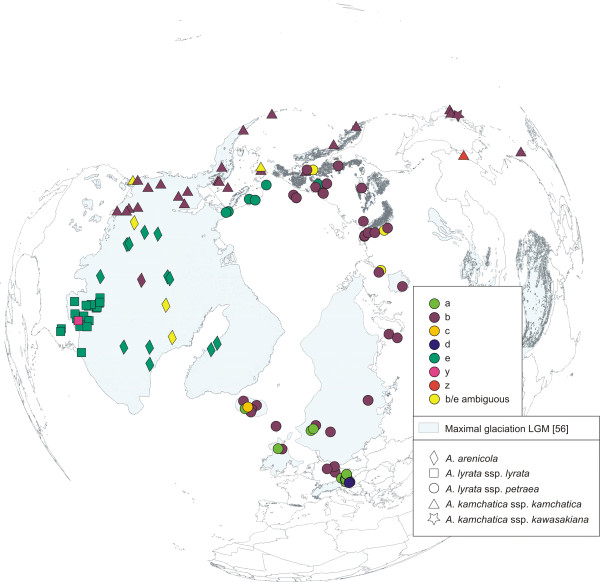
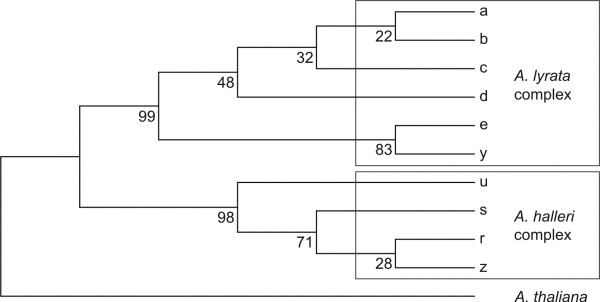
Results: Using DNA sequence data from the nuclear encoded cytosolic phosphoglucoisomerase and Internal Transcribed Spacers 1 and 2 of the ribosomal DNA, as well as the trnL/F region from the chloroplast genome, we unravelled the phylogeography of the various taxonomic units of the A. lyrata complex. We demonstrate the existence of two major gene pools in Central Europe and Northern America. These two major gene pools are constructed from different taxonomic units. We also confirmed that A. kamchatica is the allotetraploid hybrid between A. lyrata and A. halleri, occupying the amphi-Beringian area in Eastern Asia and Northern America. This species closes the large distribution gap of the various other A. lyrata segregates. Furthermore, we revealed a threefold independent allopolyploid origin of this hybrid species in Japan, China, and Kamchatka.
Conclusions: Unglaciated parts of the Eastern Austrian Alps and arctic Eurasia, including Beringia, served as major glacial refugia of the Eurasian A. lyrata lineage, whereas A. halleri and its various subspecies probably survived in refuges in Central Europe and Eastern Asia with a large distribution gap in between. The North American A. lyrata lineage probably survived the glaciation in the southeast of North America. The dramatic climatic changes during glaciation and deglaciation cycles promoted not only secondary contact and formation of the allopolyploid hybrid A. kamchatica, but also provided the environment that allowed this species to fill a large geographic gap separating the two genetically different A. lyrata lineages from Eurasia and North America. With our example focusing on the evolutionary history of the A. lyrata species complex, we add substantial information to a broad evolutionary framework for future investigations within this emerging model system in molecular and evolutionary biology.
Figures
Similar articles
-
The allopolyploid Arabidopsis kamchatica originated from multiple individuals of Arabidopsis lyrata and Arabidopsis halleri.Mol Ecol. 2009 Oct;18(19):4024-48. doi: 10.1111/j.1365-294X.2009.04329.x. Epub 2009 Sep 15. Mol Ecol. 2009. PMID: 19754506
-
Arabidopsis hybrid speciation processes.Proc Natl Acad Sci U S A. 2011 Aug 23;108(34):14192-7. doi: 10.1073/pnas.1104212108. Epub 2011 Aug 8. Proc Natl Acad Sci U S A. 2011. PMID: 21825128 Free PMC article.
-
The evolutionary history of the Arabidopsis arenosa complex: diverse tetraploids mask the Western Carpathian center of species and genetic diversity.PLoS One. 2012;7(8):e42691. doi: 10.1371/journal.pone.0042691. Epub 2012 Aug 3. PLoS One. 2012. PMID: 22880083 Free PMC article.
-
Polyploid Arabidopsis species originated around recent glaciation maxima.Curr Opin Plant Biol. 2018 Apr;42:8-15. doi: 10.1016/j.pbi.2018.01.005. Epub 2018 Feb 12. Curr Opin Plant Biol. 2018. PMID: 29448159 Review.
-
Diversity, Distribution, and Evolution of Bioluminescent Fungi.J Fungi (Basel). 2024 Dec 31;11(1):19. doi: 10.3390/jof11010019. J Fungi (Basel). 2024. PMID: 39852438 Free PMC article. Review.
Cited by
-
Multiple Autopolyploid Arabidopsis lyrata Populations Stabilized by Long-Range Adaptive Introgression Across Eurasia.Mol Biol Evol. 2025 Jul 30;42(8):msaf153. doi: 10.1093/molbev/msaf153. Mol Biol Evol. 2025. PMID: 40701836 Free PMC article.
-
Polyploid evolution and Pleistocene glacial cycles: A case study from the alpine primrose Primula marginata (Primulaceae).BMC Evol Biol. 2012 Apr 24;12:56. doi: 10.1186/1471-2148-12-56. BMC Evol Biol. 2012. PMID: 22530870 Free PMC article.
-
A Time-Calibrated Road Map of Brassicaceae Species Radiation and Evolutionary History.Plant Cell. 2015 Oct;27(10):2770-84. doi: 10.1105/tpc.15.00482. Epub 2015 Sep 26. Plant Cell. 2015. PMID: 26410304 Free PMC article.
-
Chloroplast genomes of Arabidopsis halleri ssp. gemmifera and Arabidopsis lyrata ssp. petraea: Structures and comparative analysis.Sci Rep. 2017 Aug 8;7(1):7556. doi: 10.1038/s41598-017-07891-5. Sci Rep. 2017. PMID: 28790364 Free PMC article.
-
Temperature-stress resistance and tolerance along a latitudinal cline in North American Arabidopsis lyrata.PLoS One. 2015 Jun 25;10(6):e0131808. doi: 10.1371/journal.pone.0131808. eCollection 2015. PLoS One. 2015. PMID: 26110428 Free PMC article.
References
-
- Mitchell-Olds T. Arabidopsis thaliana and its wild relatives: a model system for ecology and evolution. Trends Ecol Evol. 2001;16:693–700. doi: 10.1016/S0169-5347(01)02291-1. - DOI
-
- Koch MA, Haubold B, Mitchell-Olds T. Comparative evolutionary analysis of chalcone synthase and alcohol dehydrogenase loci in Arabidopsis, Arabis, and related genera (Brassicaceae) Mol Biol Evol. 2000;17:1483–1498. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
