

Molecular mechanism that induces activation of Spätzle, the ligand for the Drosophila Toll receptor
- PMID: 20378549
- PMCID: PMC2885229
- DOI: 10.1074/jbc.M109.098186
Molecular mechanism that induces activation of Spätzle, the ligand for the Drosophila Toll receptor
Abstract
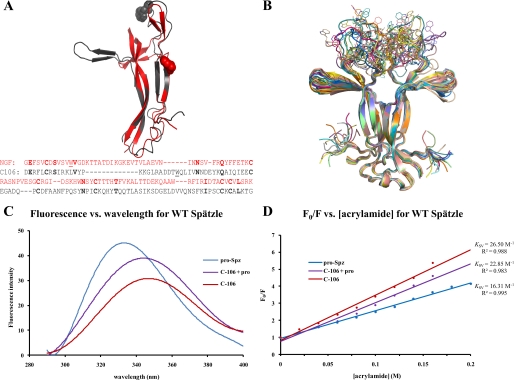
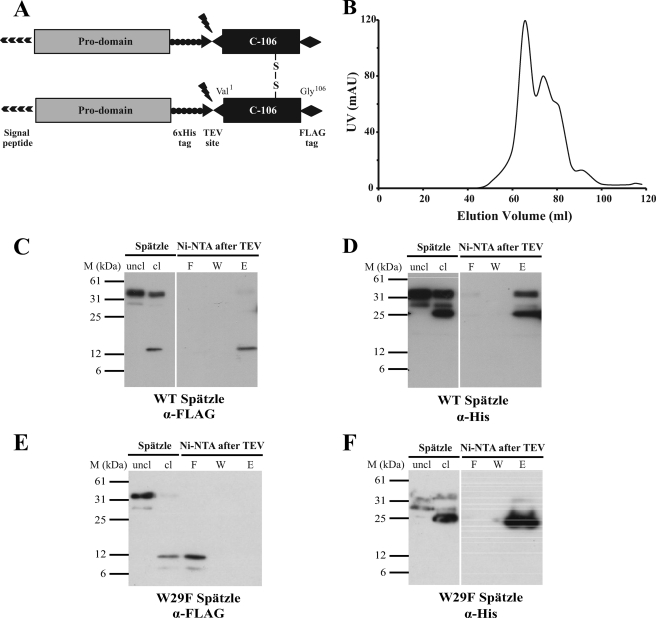
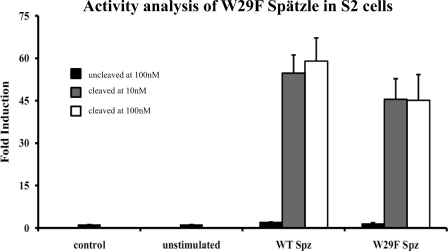
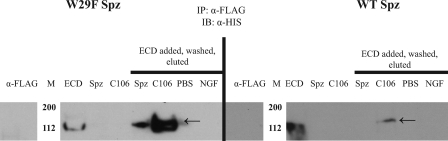
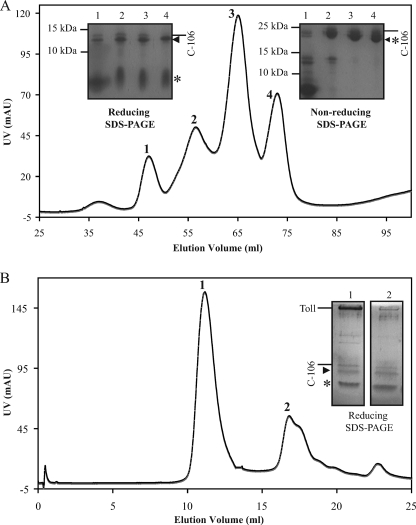
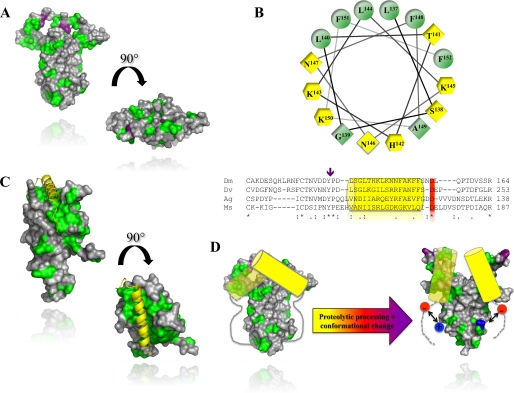
The Drosophila Toll receptor is activated by an endogenous cytokine ligand Spätzle. Active ligand is generated in response to positional cues in embryonic dorso-ventral patterning and microbial pathogens in the insect immune response. Spätzle is secreted as a pro-protein and is processed into an active form by the serine endoproteases Easter and Spätzle-processing enzyme during dorso-ventral patterning and infection, respectively. Here, we provide evidence for the molecular mechanism of this activation process. We show that the Spätzle prodomain masks a predominantly hydrophobic region of Spätzle and that proteolysis causes a conformational change that exposes determinants that are critical for binding to the Toll receptor. We also gather that a conserved sequence motif in the prodomain presents features of an amphipathic helix likely to bind a hydrophobic cleft in Spätzle thereby occluding the putative Toll binding region. This mechanism of activation has a striking similarity to that of coagulogen, a clotting factor of the horseshoe crab, an invertebrate that has changed little in 400 million years. Taken together, our findings demonstrate that an ancient passive defense system has been adapted during evolution and converted for use in a critical pathway of innate immune signaling and embryonic morphogenesis.
Figures
References
-
- Belvin M. P., Anderson K. V. (1996) Annu. Rev. Cell Dev. Biol. 12, 393–416 - PubMed
-
- Hoffmann J. A. (2003) Nature 426, 33–38 - PubMed
-
- Schneider D. S., Jin Y., Morisato D., Anderson K. V. (1994) Development 120, 1243–1250 - PubMed
-
- Weber A. N., Tauszig-Delamasure S., Hoffmann J. A., Lelièvre E., Gascan H., Ray K. P., Morse M. A., Imler J. L., Gay N. J. (2003) Nat. Immunol. 4, 794–800 - PubMed
-
- Mizuguchi K., Parker J. S., Blundell T. L., Gay N. J. (1998) Trends Biochem. Sci. 23, 239–242 - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
