

Importance of the alphaC-helix in the cyclic nucleotide binding domain for the stable channel regulation and function of cyclic nucleotide gated ion channels in Arabidopsis
- PMID: 20378667
- PMCID: PMC2877894
- DOI: 10.1093/jxb/erq072
Importance of the alphaC-helix in the cyclic nucleotide binding domain for the stable channel regulation and function of cyclic nucleotide gated ion channels in Arabidopsis
Abstract
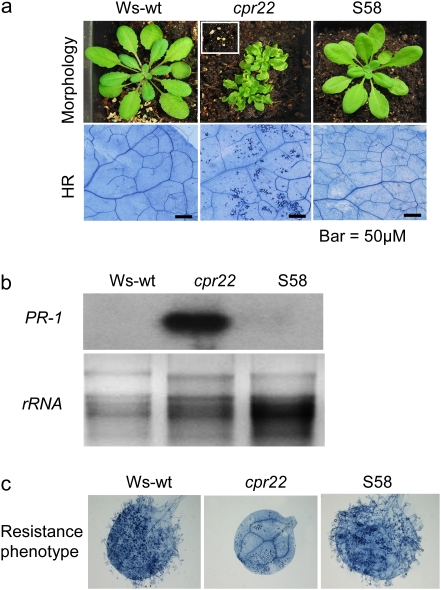
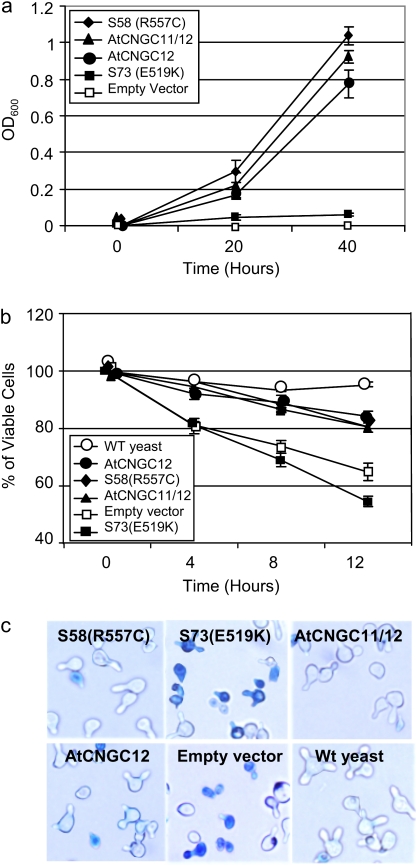
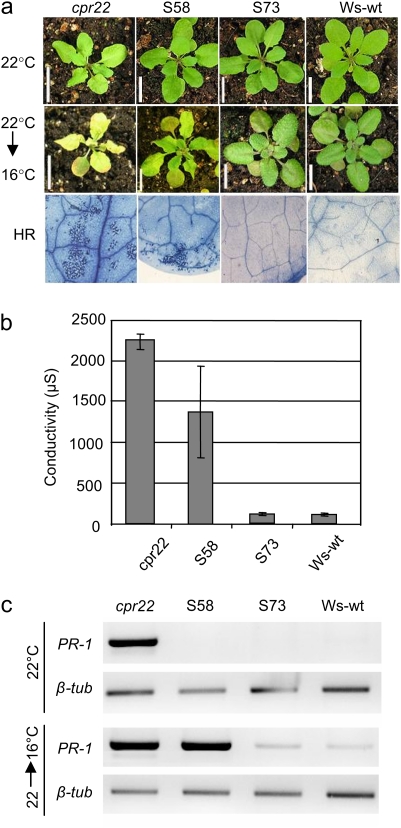
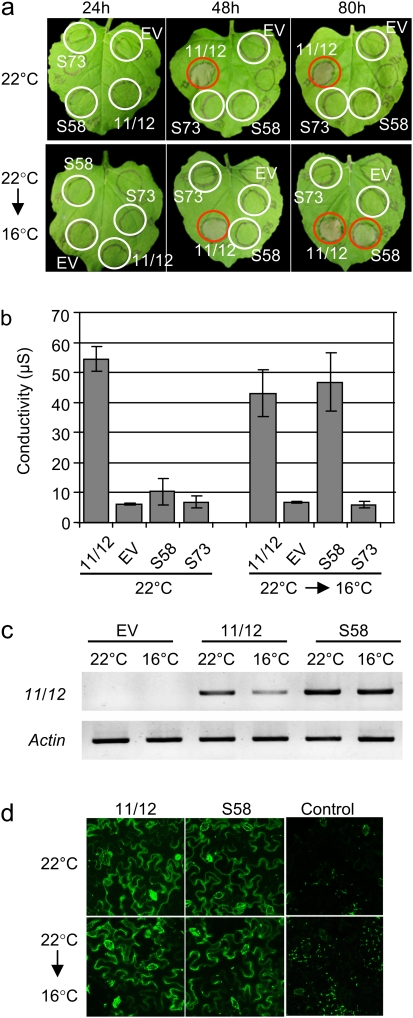
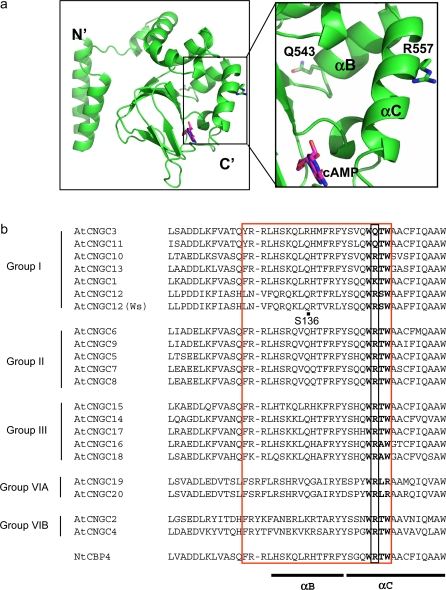
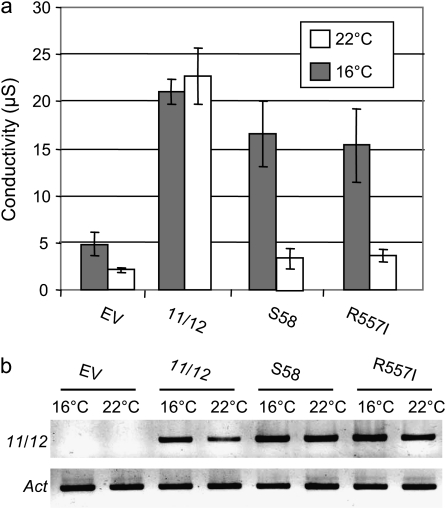
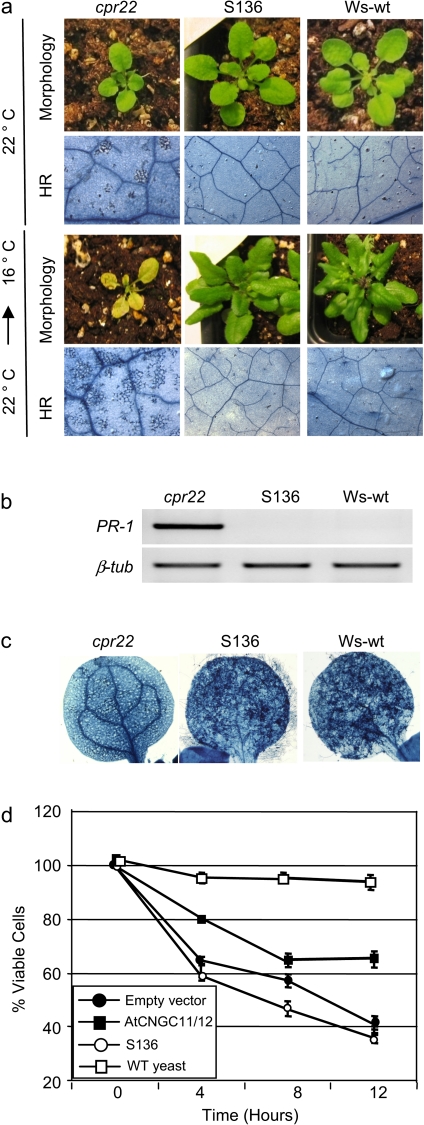
The involvement of cyclic nucleotide gated ion channels (CNGCs) in the signal transduction of animal light and odorant perception is well documented. Although plant CNGCs have recently been revealed to mediate multiple stress responses and developmental pathways, studies that aim to elucidate their structural and regulatory properties are still very much in their infancy. The structure-function relationship of plant CNGCs was investigated here by using the chimeric Arabidopsis AtCNGC11/12 gene that induces multiple defence responses in the Arabidopsis mutant constitutive expresser of PR genes 22 (cpr22) for the identification of functionally essential residues. A genetic screen for mutants that suppress cpr22-conferred phenotypes identified over 20 novel mutant alleles in AtCNGC11/12. One of these mutants, suppressor S58 possesses a single amino acid substitution, arginine 557 to cysteine, in the alphaC-helix of the cyclic nucleotide-binding domain (CNBD). The suppressor S58 lost all cpr22 related phenotypes, such as spontaneous cell death formation under ambient temperature conditions. However, these phenotypes were recovered at 16 degrees C suggesting that the stability of channel function is affected by temperature. In silico modelling and site-directed mutagenesis analyses suggest that arginine 557 in the alphaC-helix of the CNBD is important for channel regulation, but not for basic function. Furthermore, another suppressor mutant, S136 that lacks the entire alphaC-helix due to a premature stop codon, lost channel function completely. Our data presented here indicate that the alphaC-helix is functionally important in plant CNGCs.
Figures
References
-
- Ali R, Zielinski RE, Berkowitz GA. Expression of plant cyclic nucleotide-gated cation channels in yeast. Journal of Experimental Botany. 2006;57:125–138. - PubMed
-
- Arazi T, Sunkar R, Kaplan B, Fromm H. A tobacco plasma membrane calmodulin-binding transporter confers Ni2+ tolerance and Pb2+ hypersensitivity in transgenic plants. The Plant Journal. 1999;20:171–182. - PubMed
-
- Arazi T, Kaplan B, Fromm H. A high-affinity calmodulin-binding site in tobacco plasma-membrane channel protein coincides with a characteristic element of cyclic nucleotide-binding domains. Plant Molecular Biology. 2000;42:591–601. - PubMed
-
- Ausubel FM, Bent R, Kingston RE, Moore DD, Seidman JG, Smith JA, Struhl K. Current protocols in molecular biology. New York: John Wiley and Sons; 1987.
-
- Baxter J, Moeder W, Urquhart W, Shahinas D, Chin K, Christendat D, Kang HG, Angelova M, Kato N, Yoshioka K. Identification of a functionally essential amino acid for Arabidopsis cyclic nucleotide gated ion channels using the chimeric AtCNGC11/12 gene. The Plant Journal. 2008;56:457–469. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
