

Targeting X chromosomes for repression
- PMID: 20381335
- PMCID: PMC2906122
- DOI: 10.1016/j.gde.2010.03.008
Targeting X chromosomes for repression
Abstract
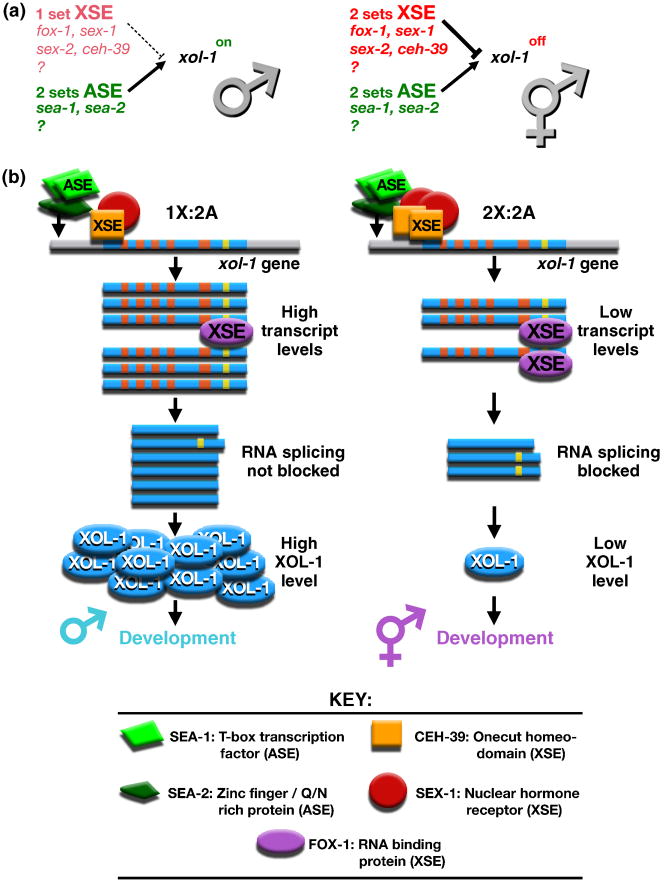
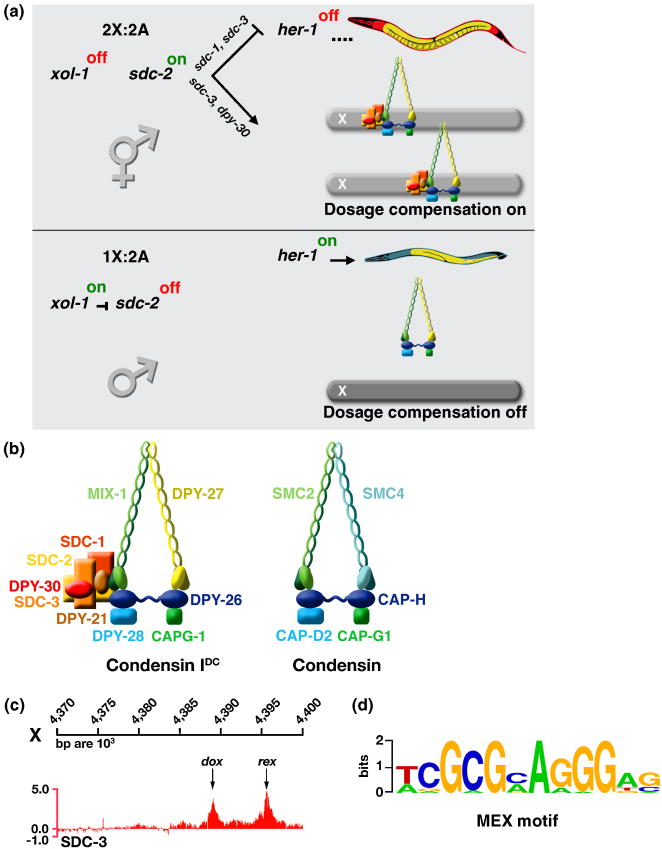
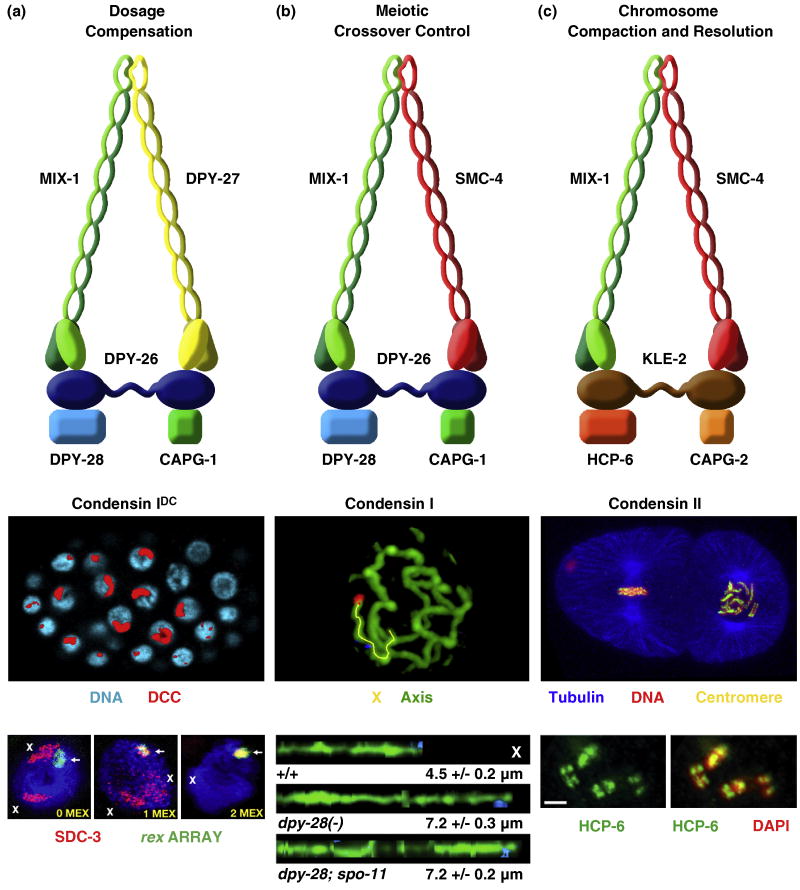
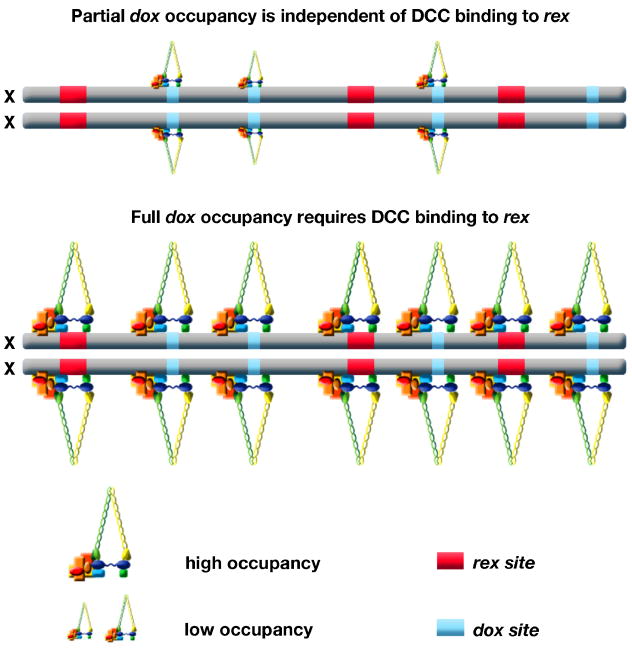
Dosage compensation is a chromosome-wide regulatory process that balances X-chromosome gene expression between males and females that have different complements. Recent advances have clarified the molecular nature of the Caenorhabditis elegans sex-determination signal, which tallies X-chromosome number relative to the ploidy and controls both the choice of sexual fate and the process of dosage compensation. Dissecting the sex signal has revealed molecular mechanisms by which small quantitative differences in intracellular signals are translated into dramatically different developmental fates. Recent experiments have also revealed fundamental principles by which C. elegans dosage compensation proteins recognize and bind X chromosomes of XX embryos to reduce gene expression. Dosage compensation proteins function not only in a condensin complex specialized for regulating X-chromosome gene expression, but also in distinct condensin complexes that control other chromosome-wide processes: chromosome segregation and meiotic crossover recombination. The reshuffling of interchangeable molecular parts creates independent machines with similar architecture but distinct biological functions.
2010 Elsevier Ltd. All rights reserved.
Figures
References
-
- Charlesworth B. The evolution of chromosomal sex determination and dosage compensation. Curr Biol. 1996;6:149–162. - PubMed
-
- Payer B, Lee JT. X chromosome dosage compensation: how mammals keep the balance. Annu Rev Genet. 2008;42:733–772. - PubMed
-
- Bacher CP, Guggiari M, Brors B, Augui S, Clerc P, Avner P, Eils R, Heard E. Transient colocalization of X-inactivation centres accompanies the initiation of X inactivation. Nat Cell Biol. 2006;(8):293–299. - PubMed
-
- Xu N, Tsai CL, Lee JT. Transient homologous chromosome pairing marks the onset of X inactivation. Science. 2006;311:1149–1152. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
