

Inhibition of electron transport chain assembly and function promotes photodynamic killing of Candida
- PMID: 20381373
- PMCID: PMC2881298
- DOI: 10.1016/j.jphotobiol.2010.03.005
Inhibition of electron transport chain assembly and function promotes photodynamic killing of Candida
Abstract
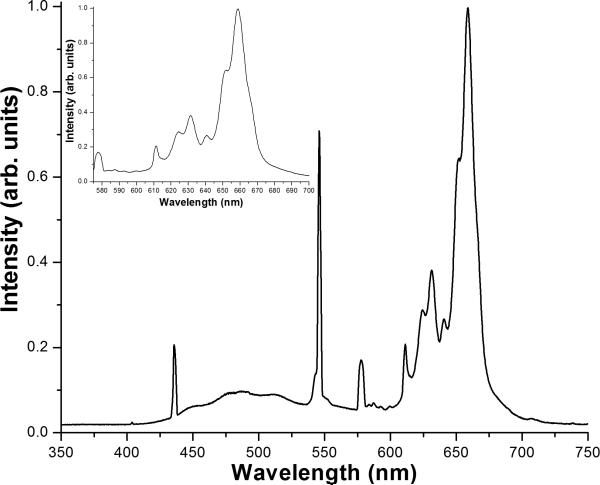
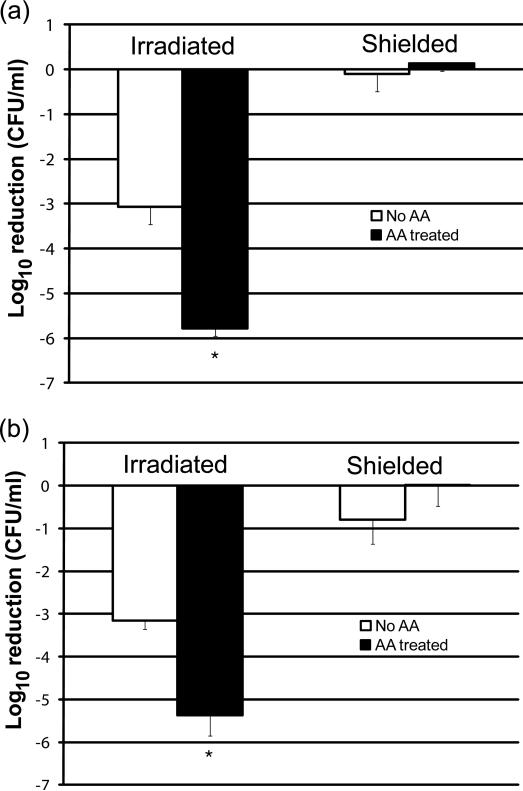
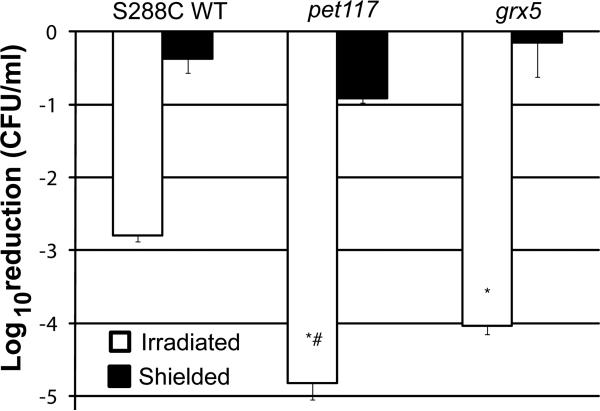
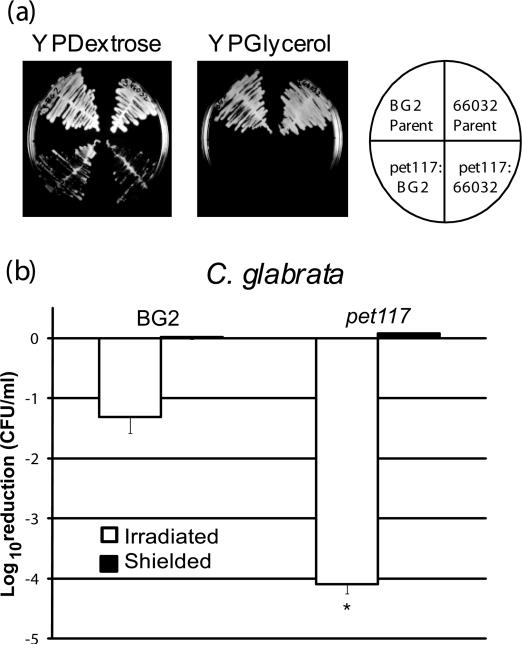
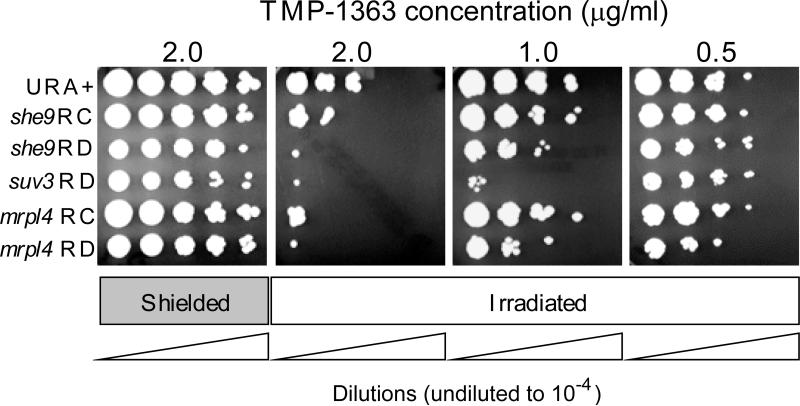
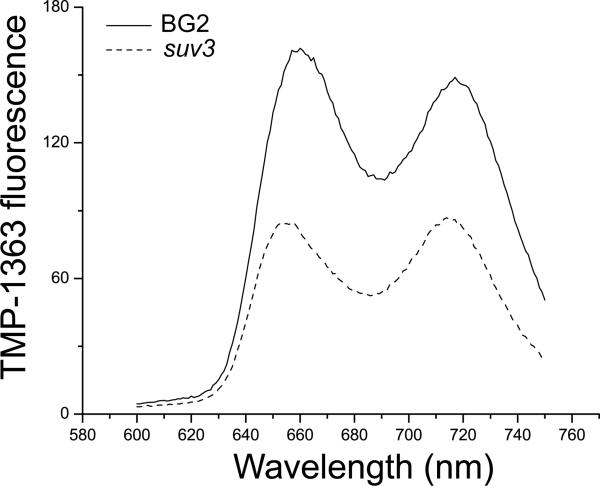
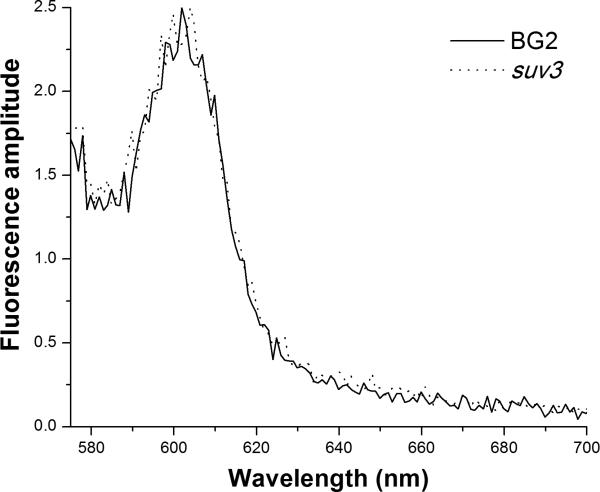
Respiratory deficiency increases the sensitivity of the pathogenic fungi Candida albicans and Candida glabrata to oxidative stress induced by photodynamic therapy (PDT) sensitized by the cationic porphyrin meso-tetra (N-methyl-4-pyridyl) porphine tetra tosylate (TMP-1363). Since disruption of electron transport chain (ETC) function increases intracellular levels of reactive oxygen species in yeast, we determined whether interference with ETC assembly or function increased sensitivity to TMP-1363-PDT in C. albicans, C. glabrata and the non-pathogenic yeast Saccharomyces cerevisiae. Metabolic inhibitor antimycin A and defined genetic mutants were used to identify ETC components that contribute to the sensitivity to PDT. Inhibition of cytochrome bc(1) (Complex III) with antimycin A increases mitochondrial levels of reactive oxygen species. PDT performed following pre-treatment with antimycin A reduced colony forming units (CFU) of C. albicans and C. glabrata by approximately two orders of magnitude relative to PDT alone. A S. cerevisiae mitochondrial glutaredoxin grx5 mutant, defective in assembly of Fe-S clusters critical for Complex III function, displayed increased sensitivity to PDT. Furthermore, C. glabrata and S.cerevisiae mutants in cytochrome c oxidase (Complex IV) synthesis and assembly were also significantly more sensitive to PDT. These included suv3, encoding an ATP-dependent RNA helicase critical for maturation of cytochrome c oxidase subunit transcripts, and pet117, encoding an essential cytochrome c oxidase assembly factor. Following PDT, the reduction in CFU of these mutants was one to two orders of magnitude greater than in their respective parental strains. The data demonstrate that selective inhibition of ETC Complexes III and IV significantly increases the sensitivity of C. albicans, C. glabrata and S. cerevisiae to PDT sensitized with TMP-1363.
2010 Elsevier B.V. All rights reserved.
Figures
References
-
- Calderone RA. Candida and Candidiasis. ASM Press; Washington, D.C.: 2002.
-
- Pfaller MA, Messer SA, Hollis RJ, Jones RN, Doern GV, Brandt ME, Hajjeh RA. Trends in species distribution and susceptibility to fluconazole among blood stream isolates of Candida species in the United States. Diag. Microbiol. Infect. Dis. 1999;33:217–222. - PubMed
-
- Johnson EM, Warnock DW, Luker J, Porter SR, Scully C. Emergence of azole drug resistance in Candida species from HIV-infected patients receiving prolonged fluconazole therapy for oral candidosis. J. Antimicrob. Chemother. 1995;35:103–114. - PubMed
-
- Li L, Redding S, Dongari-Bagtzoglou A. Candida glabrata: an emerging oral opportunistic pathogen. J. Dent. Res. 2007;86:204–215. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
