

Rescue and repair during photoreceptor cell renewal mediated by docosahexaenoic acid-derived neuroprotectin D1
- PMID: 20382842
- PMCID: PMC2903812
- DOI: 10.1194/jlr.R001131
Rescue and repair during photoreceptor cell renewal mediated by docosahexaenoic acid-derived neuroprotectin D1
Abstract
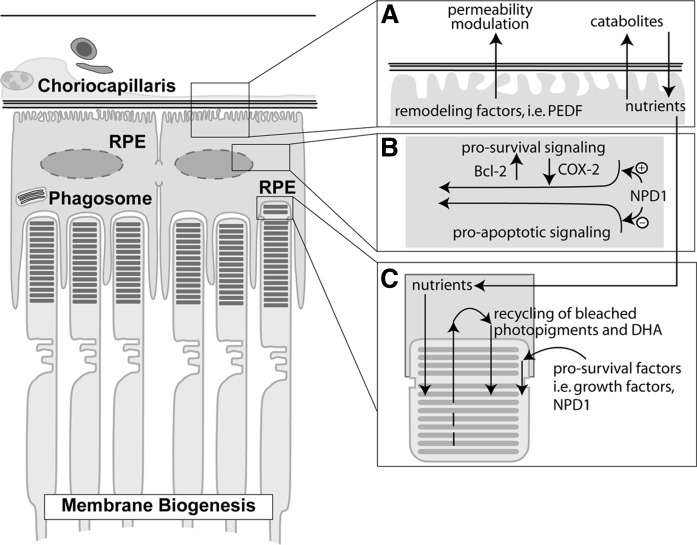
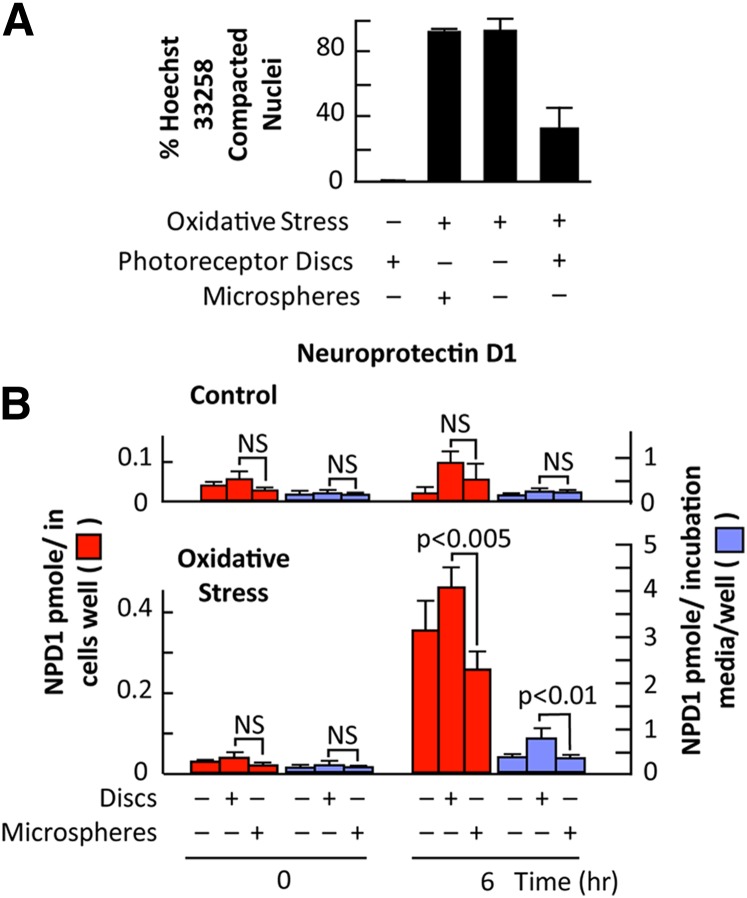
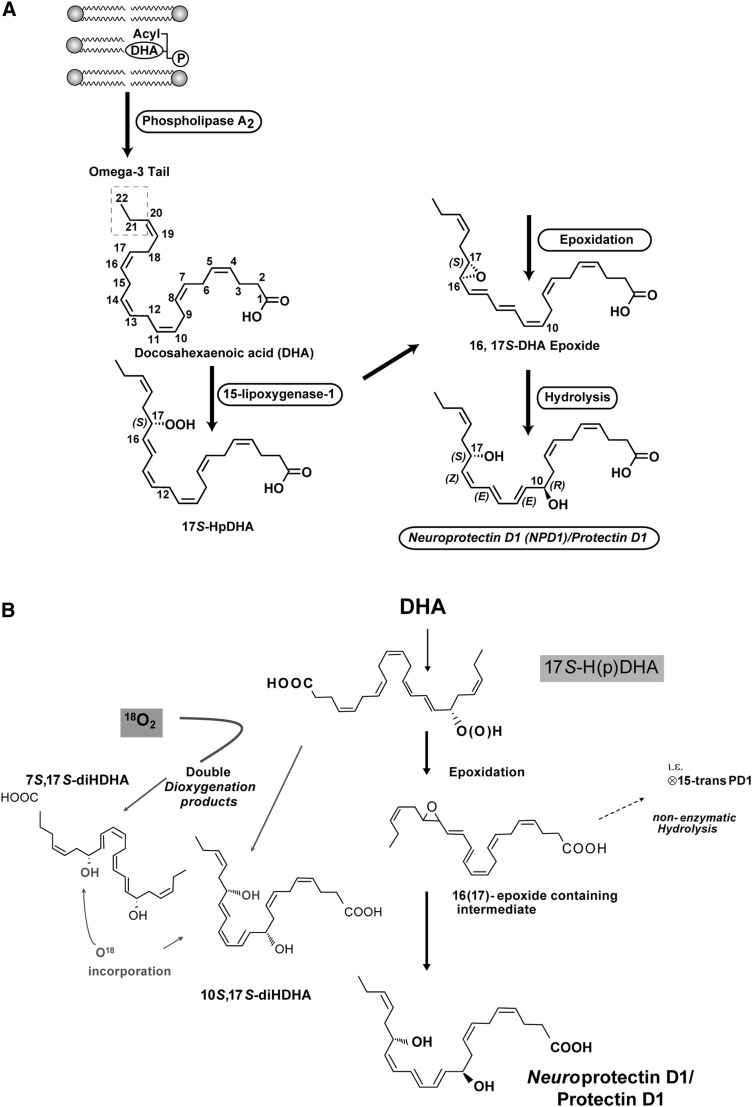
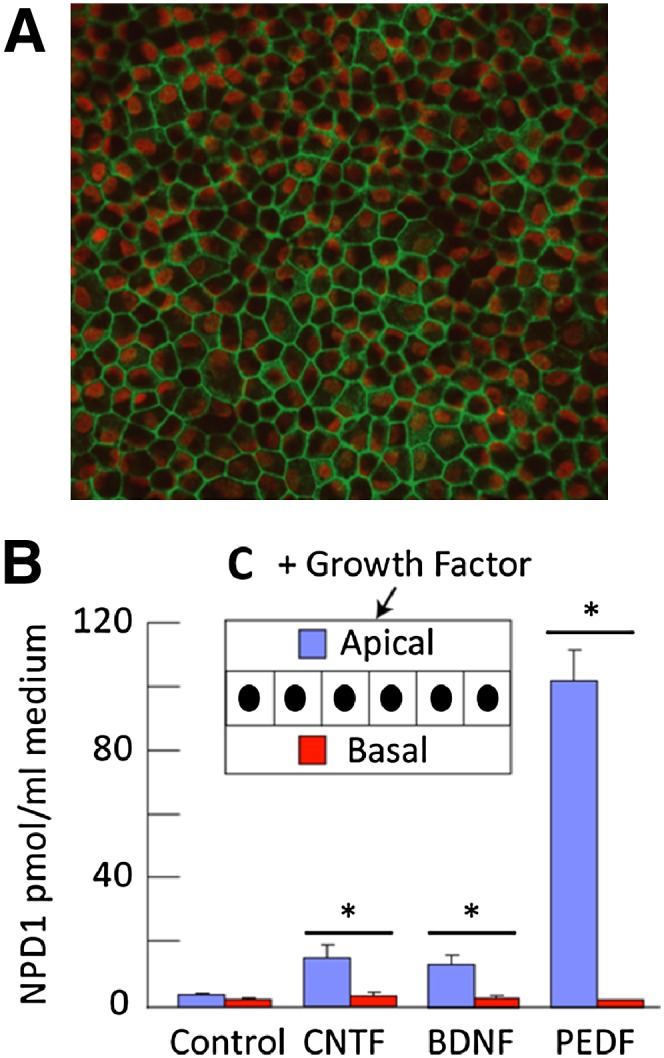
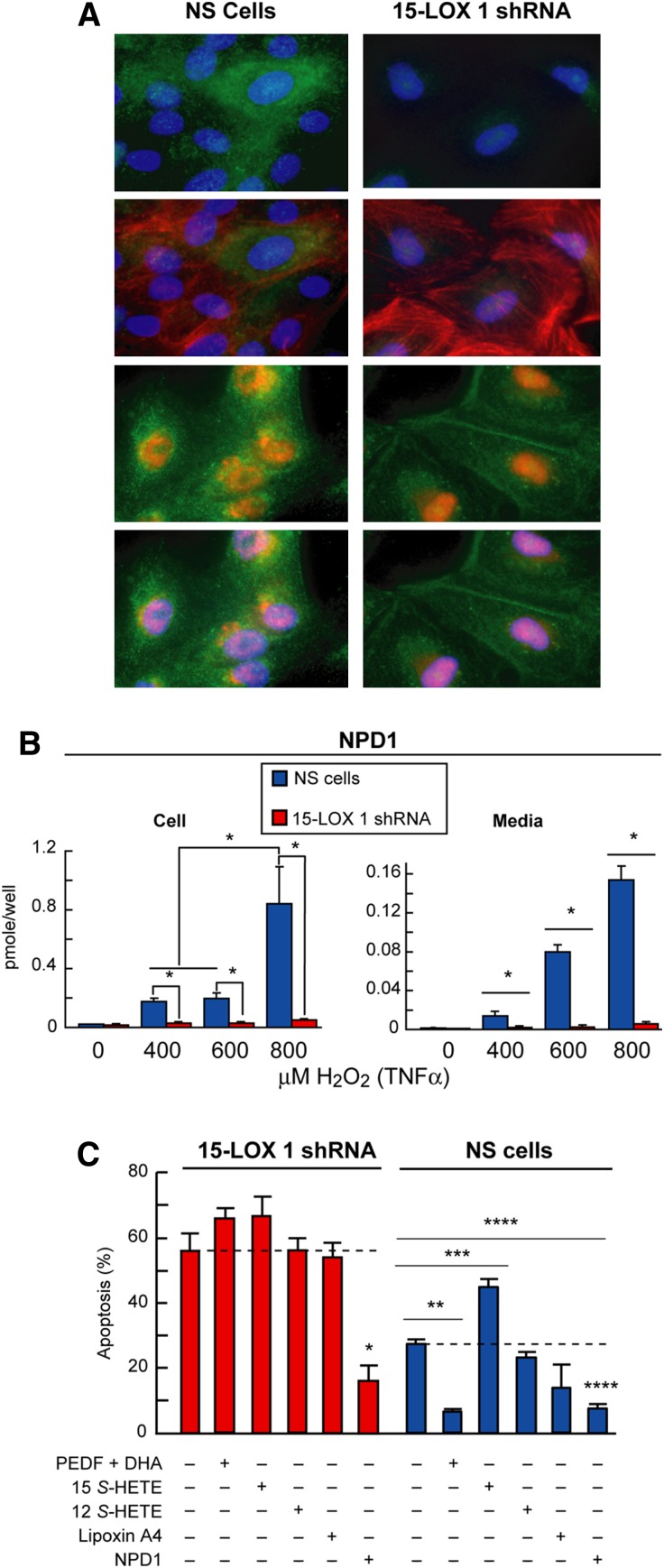
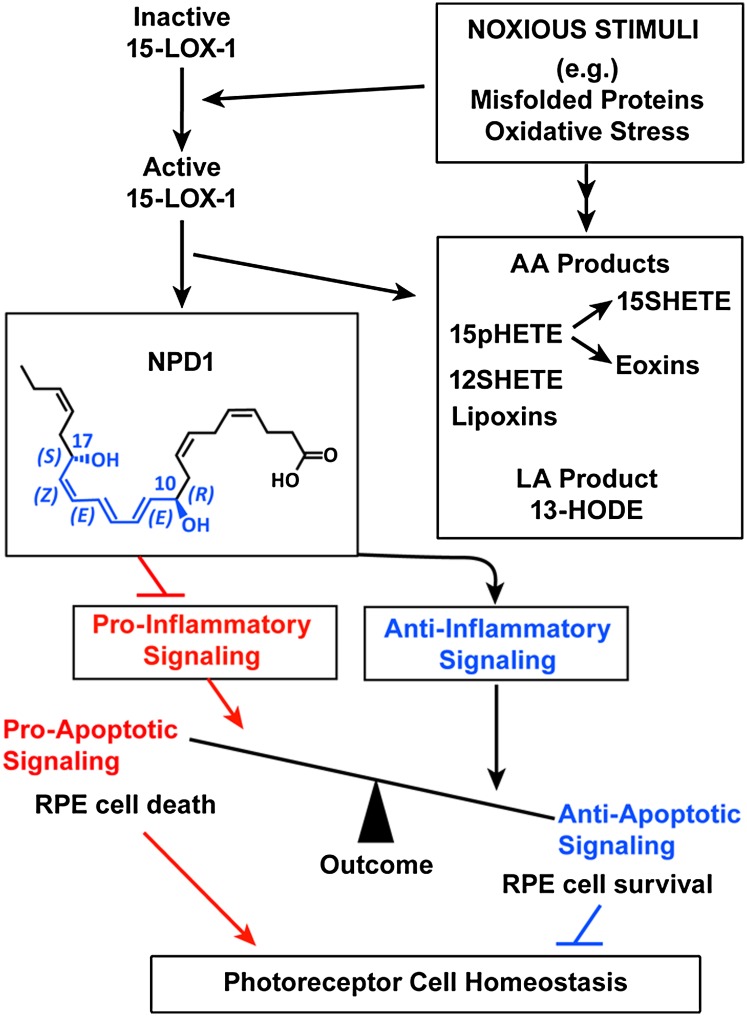
Retinal degenerative diseases result in retinal pigment epithelial (RPE) and photoreceptor cell loss. These cells are continuously exposed to the environment (light) and to potentially pro-oxidative conditions, as the retina's oxygen consumption is very high. There is also a high flux of docosahexaenoic acid (DHA), a PUFA that moves through the blood stream toward photoreceptors and between them and RPE cells. Photoreceptor outer segment shedding and phagocytosis intermittently renews photoreceptor membranes. DHA is converted through 15-lipoxygenase-1 into neuroprotectin D1 (NPD1), a potent mediator that evokes counteracting cell-protective, anti-inflammatory, pro-survival repair signaling, including the induction of anti-apoptotic proteins and inhibition of pro-apoptotic proteins. Thus, NPD1 triggers activation of signaling pathway/s that modulate/s pro-apoptotic signals, promoting cell survival. This review provides an overview of DHA in photoreceptors and describes the ability of RPE cells to synthesize NPD1 from DHA. It also describes the role of neurotrophins as agonists of NPD1 synthesis and how photoreceptor phagocytosis induces refractoriness to oxidative stress in RPE cells, with concomitant NPD1 synthesis.
Figures
References
-
- Papermaster D. S. 2002. The birth and death of photoreceptors: the Friedenwald Lecture. Invest. Ophthalmol. Vis. Sci. 43: 1300–1309. - PubMed
-
- Rattner A., Nathans J. 2006. Macular degeneration: recent advances and therapeutic opportunities. Nat. Rev. Neurosci. 7: 860–872. - PubMed
-
- Chang G. Q., Hao Y., Wong F. 1993. Apoptosis: final common pathway of photoreceptor death in rd, rds, and rhodopsin mutant mice. Neuron. 11: 595–605. - PubMed
-
- Bird A. C. 2003. The Bowman lecture. Towards an understanding of age-related macular disease. Eye (Lond.). 17: 457–466. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
