

Inositol trisphosphate receptor Ca2+ release channels in neurological diseases
- PMID: 20383523
- PMCID: PMC2893360
- DOI: 10.1007/s00424-010-0826-0
Inositol trisphosphate receptor Ca2+ release channels in neurological diseases
Abstract
The modulation of cytoplasmic Ca2+ concentration by release from internal stores through the inositol trisphosphate receptor (InsP3R) Ca2+ release channel is a ubiquitous signaling system involved in the regulation of numerous processes. Because of its ubiquitous expression and roles in regulating diverse cell physiological processes, it is not surprising that the InsP3R has been implicated in a number of disease states. However, relatively few mutations in InsP3R genes have been identified to date. Here, I will discuss mutations in the type 1 InsP3R that have been discovered by analyses of human patients and mice with neurological disorders. In addition, I will highlight diseases caused by mutations in other genes, including Huntington's and Alzheimer's diseases and some spinocerebellar ataxias, where the mutant proteins have been found to exert strong influences on InsP3R function that may link InsP3R to disease pathogenesis.
Figures
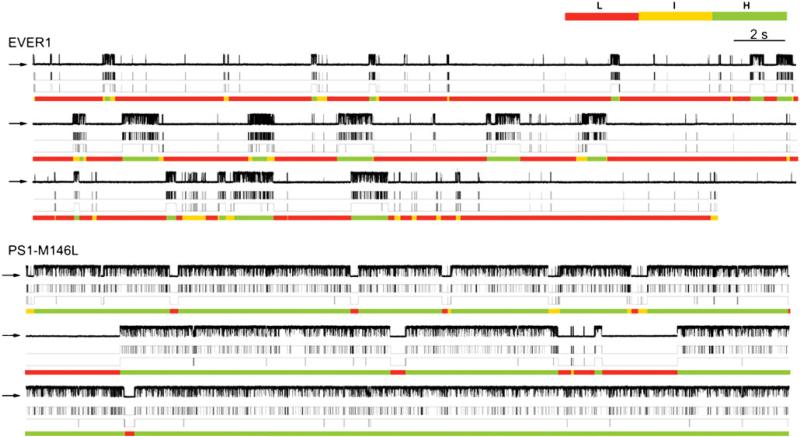
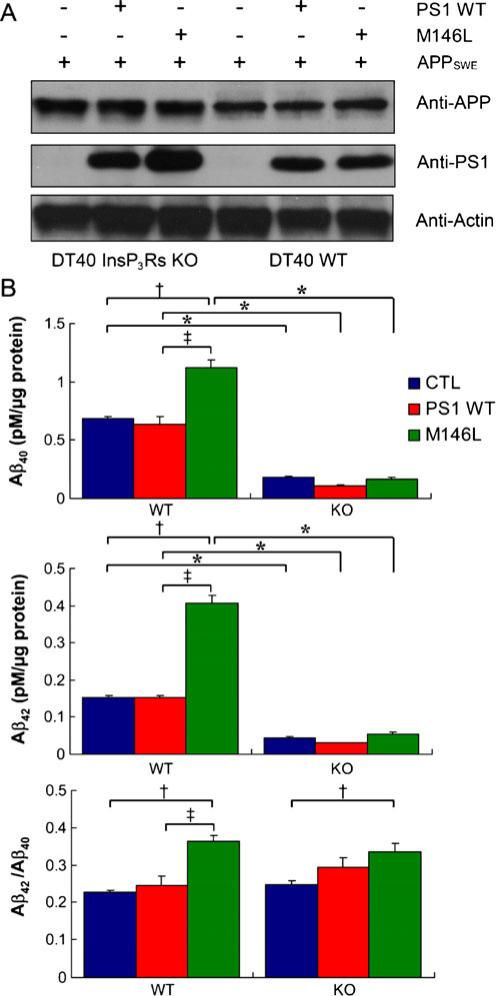
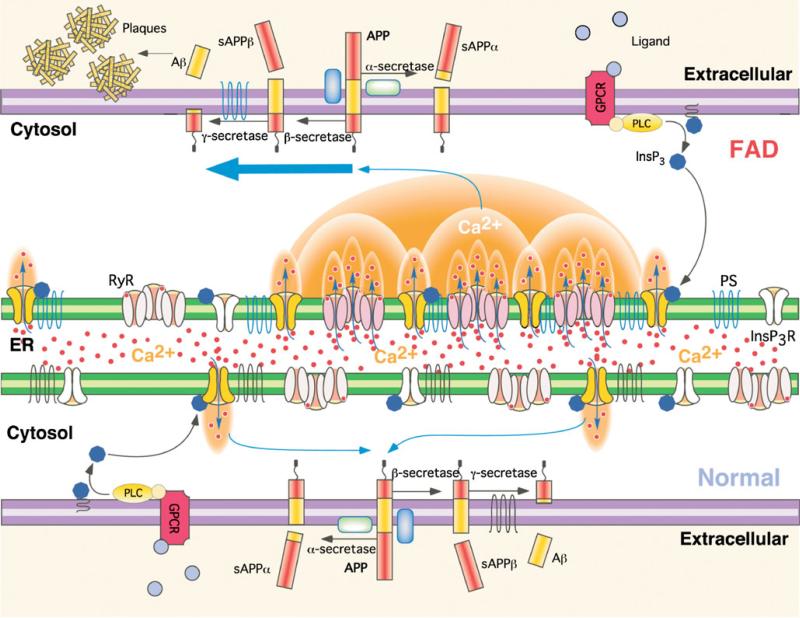
References
-
- Ando H, Mizutani A, Matsu-ura T, Mikoshiba K. IRBIT, a novel inositol 1,4,5-trisphosphate (IP3) receptor-binding protein, is released from the IP3 receptor upon IP3 binding to the receptor. J Biol Chem. 2003;278:10602–10612. - PubMed
-
- Berridge MJ, Bootman MD, Lipp P. Calcium—a life and death signal. Nature. 1998;395:645–648. - PubMed
-
- Boehning D, Mak D-OD, Foskett JK, Joseph SK. Molecular determinants of ion permeation and selectivity in inositol 1,4,5-trisphosphate receptor Ca2+ channels. J Biol Chem. 2001;276:13509–13512. - PubMed
-
- Bosanac I, Alattia JR, Mal TK, Chan J, Talarico S, Tong FK, Tong KI, Yoshikawa F, Furuichi T, Iwai M, Michikawa T, Mikoshiba K, Ikura M. Structure of the inositol 1,4,5-trisphosphate receptor binding core in complex with its ligand. Nature. 2002;420:696–700. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
