

Growth of collagen fibril seeds from embryonic tendon: fractured fibril ends nucleate new tip growth
- PMID: 20385142
- PMCID: PMC2887488
- DOI: 10.1016/j.jmb.2010.04.008
Growth of collagen fibril seeds from embryonic tendon: fractured fibril ends nucleate new tip growth
Abstract
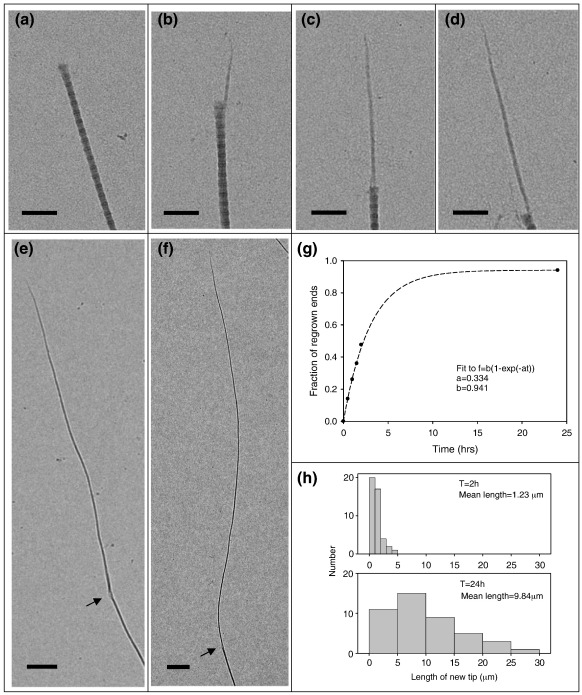
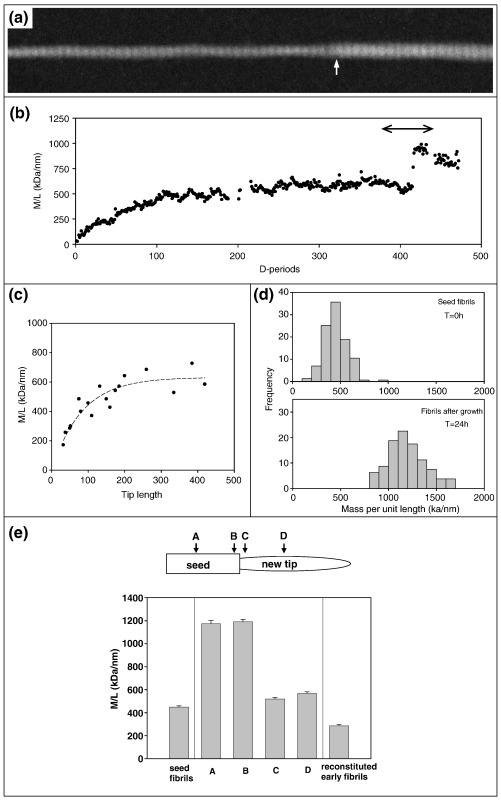
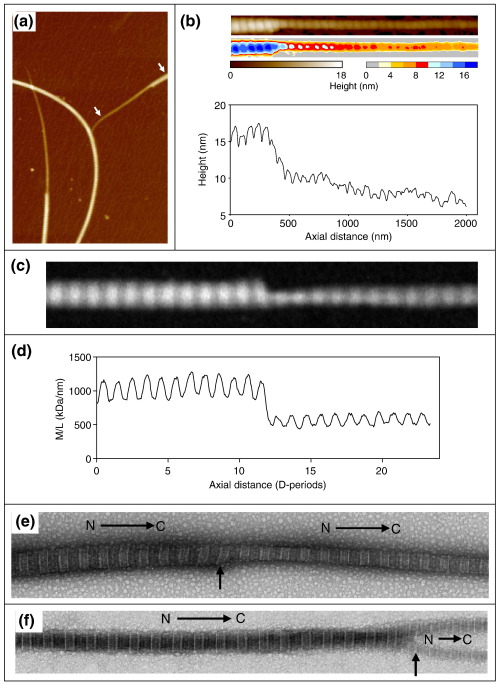
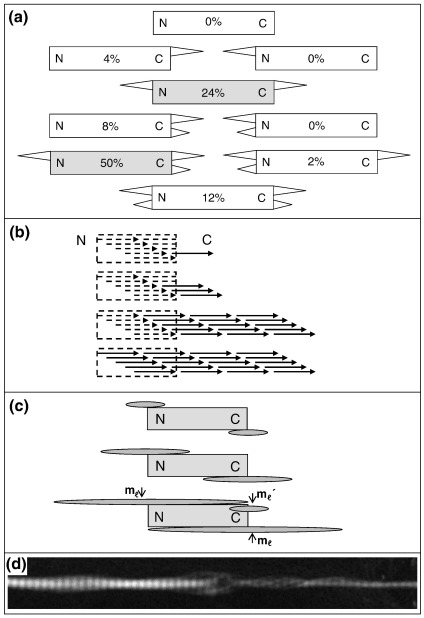
Collagen fibrils are the principal tensile element of vertebrate tissues where they occur in the extracellular matrix as spatially organised arrays. A major challenge is to understand how the mechanisms of nucleation, growth and remodelling yield fibrils of tissue-specific diameter and length. Here we have developed a seeding system whereby collagen fibrils were isolated from avian embryonic tendon and added to purified collagen solution, in order to characterise fibril surface nucleation and growth mechanisms. Fragmentation of tendon in liquid nitrogen followed by Dounce homogenisation generated fibril length fragments. Most (>94%) of the fractured ends of fibrils, which show an abrupt square profile, were found to act as nucleation sites for further growth by molecular accretion. The mechanism of this nucleation and growth process was investigated by transmission electron microscopy, atomic force microscopy and scanning transmission electron microscopy mass mapping. Typically, a single growth spur occurred on the N-terminal end of seed fibrils whilst twin spurs frequently formed on the C-terminal end before merging into a single tip projection. The surface nucleation and growth process generated a smoothly tapered tip that achieved maximum diameter when the axial extension reached approximately 13 mum. Lateral growth also occurred along the entire length of all seed fibrils that contained tip projections. The data support a model of collagen fibril growth in which the broken ends of fibrils are nucleation sites for propagation in opposite axial directions. The observed fibril growth behaviour has direct relevance to tendon matrix remodelling and repair processes that might involve rupture of collagen fibrils.
Copyright (c) 2010 Elsevier Ltd. All rights reserved.
Figures
Similar articles
-
Collagen fibrils forming in developing tendon show an early and abrupt limitation in diameter at the growing tips.J Mol Biol. 1998 Nov 13;283(5):1049-58. doi: 10.1006/jmbi.1998.2153. J Mol Biol. 1998. PMID: 9799643
-
Identification of collagen fibril fusion during vertebrate tendon morphogenesis. The process relies on unipolar fibrils and is regulated by collagen-proteoglycan interaction.J Mol Biol. 2000 Jan 28;295(4):891-902. doi: 10.1006/jmbi.1999.3384. J Mol Biol. 2000. PMID: 10656798
-
A structure-based extracellular matrix expansion mechanism of fibrous tissue growth.Elife. 2015 May 20;4:e05958. doi: 10.7554/eLife.05958. Elife. 2015. PMID: 25992598 Free PMC article.
-
STEM/TEM studies of collagen fibril assembly.Micron. 2001 Apr;32(3):273-85. doi: 10.1016/s0968-4328(00)00040-8. Micron. 2001. PMID: 11006507 Review.
-
Tip-mediated fusion involving unipolar collagen fibrils accounts for rapid fibril elongation, the occurrence of fibrillar branched networks in skin and the paucity of collagen fibril ends in vertebrates.Matrix Biol. 2000 Aug;19(4):359-65. doi: 10.1016/s0945-053x(00)00082-2. Matrix Biol. 2000. PMID: 10963997 Review.
Cited by
-
Adaptive Remodeling of Achilles Tendon: A Multi-scale Computational Model.PLoS Comput Biol. 2016 Sep 29;12(9):e1005106. doi: 10.1371/journal.pcbi.1005106. eCollection 2016 Sep. PLoS Comput Biol. 2016. PMID: 27684554 Free PMC article.
-
Collagen Hybridizing Peptides Promote Collagen Fibril Growth In Vitro.ACS Appl Bio Mater. 2025 Mar 17;8(3):2003-2014. doi: 10.1021/acsabm.4c01509. Epub 2025 Feb 26. ACS Appl Bio Mater. 2025. PMID: 40010706 Free PMC article.
-
Collagenous Extracellular Matrix Biomaterials for Tissue Engineering: Lessons from the Common Sea Urchin Tissue.Int J Mol Sci. 2017 Apr 25;18(5):901. doi: 10.3390/ijms18050901. Int J Mol Sci. 2017. PMID: 28441344 Free PMC article. Review.
-
Tendon Extracellular Matrix Assembly, Maintenance and Dysregulation Throughout Life.Adv Exp Med Biol. 2021;1348:45-103. doi: 10.1007/978-3-030-80614-9_3. Adv Exp Med Biol. 2021. PMID: 34807415 Review.
-
Using tools in mechanobiology to repair tendons.Curr Tissue Microenviron Rep. 2020 Jun;1(2):31-40. doi: 10.1007/s43152-020-00005-w. Epub 2021 Mar 31. Curr Tissue Microenviron Rep. 2020. PMID: 33585822 Free PMC article.
References
-
- Fitton-Jackson S. The development of avian tendon. Proc. R. Soc. London, Ser. B. 1956;144:556–572. - PubMed
-
- Birk D.E., Nurminskaya M.V., Zycband E.I. Collagen fibrillogenesis in situ: fibril segments undergo post-depositional modifications resulting in linear and lateral growth during matrix development. Dev. Dyn. 1995;202:229–243. - PubMed
-
- Parry D.A., Barnes G.R., Craig A.S. A comparison of the size distribution of collagen fibrils in connective tissues as a function of age and a possible relation between fibril size distribution and mechanical properties. Proc. R. Soc. London, Ser. B. 1978;203:305–321. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
