

Structural basis of chaperone recognition of type III secretion system minor translocator proteins
- PMID: 20385547
- PMCID: PMC2906315
- DOI: 10.1074/jbc.M110.111278
Structural basis of chaperone recognition of type III secretion system minor translocator proteins
Abstract
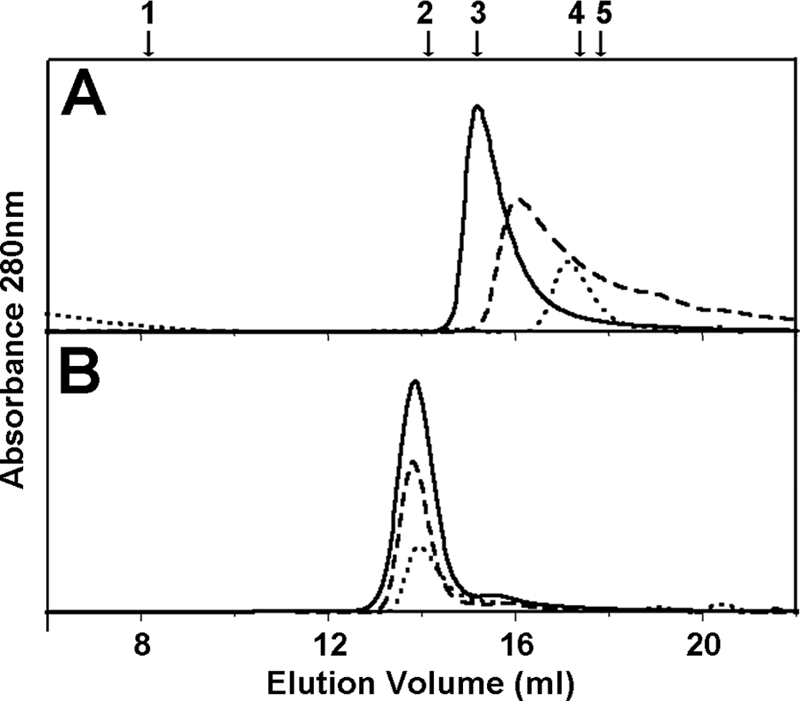
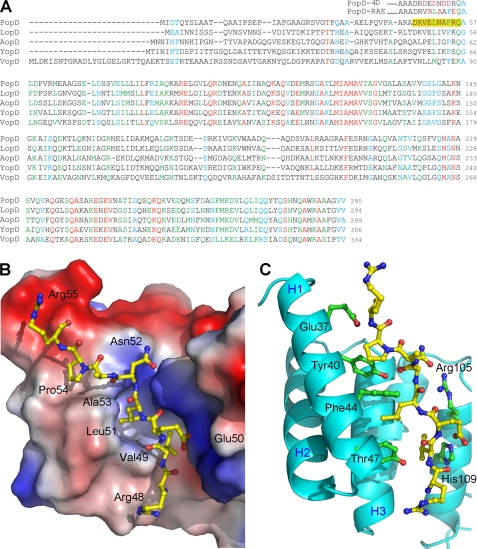
The type III secretion system (T3SS) is a complex nanomachine employed by many Gram-negative pathogens, including the nosocomial agent Pseudomonas aeruginosa, to inject toxins directly into the cytoplasm of eukaryotic cells. A key component of all T3SS is the translocon, a proteinaceous channel that is inserted into the target membrane, which allows passage of toxins into target cells. In most bacterial species, two distinct membrane proteins (the "translocators") are involved in translocon formation, whereas in the bacterial cytoplasm, however, they remain associated to a common chaperone. To date, the strategy employed by a single chaperone to recognize two distinct translocators is unknown. Here, we report the crystal structure of a complex between the Pseudomonas translocator chaperone PcrH and a short region from the minor translocator PopD. PcrH displays a 7-helical tetratricopeptide repeat fold that harbors the PopD peptide within its concave region, originally believed to be involved in recognition of the major translocator, PopB. Point mutations introduced into the PcrH-interacting region of PopD impede translocator-chaperone recognition in vitro and lead to impairment of bacterial cytotoxicity toward macrophages in vivo. These results indicate that T3SS translocator chaperones form binary complexes with their partner molecules, and the stability of their interaction regions must be strictly maintained to guarantee bacterial infectivity. The PcrH-PopD complex displays homologs among a number of pathogenic strains and could represent a novel, potential target for antibiotic development.
Figures



Similar articles
-
Membrane and chaperone recognition by the major translocator protein PopB of the type III secretion system of Pseudomonas aeruginosa.J Biol Chem. 2014 Feb 7;289(6):3591-601. doi: 10.1074/jbc.M113.517920. Epub 2013 Dec 2. J Biol Chem. 2014. PMID: 24297169 Free PMC article.
-
Binding mode analysis of a major T3SS translocator protein PopB with its chaperone PcrH from Pseudomonas aeruginosa.Proteins. 2014 Dec;82(12):3273-85. doi: 10.1002/prot.24666. Epub 2014 Oct 21. Proteins. 2014. PMID: 25116453
-
Crystal structure of the Yersinia enterocolitica type III secretion chaperone SycD in complex with a peptide of the minor translocator YopD.BMC Struct Biol. 2012 Jun 18;12:13. doi: 10.1186/1472-6807-12-13. BMC Struct Biol. 2012. PMID: 22708907 Free PMC article.
-
Membrane targeting and pore formation by the type III secretion system translocon.FEBS J. 2011 Feb;278(3):414-26. doi: 10.1111/j.1742-4658.2010.07974.x. Epub 2010 Dec 23. FEBS J. 2011. PMID: 21182592 Review.
-
Key steps in type III secretion system (T3SS) towards translocon assembly with potential sensor at plant plasma membrane.Mol Plant Pathol. 2015 Sep;16(7):762-73. doi: 10.1111/mpp.12223. Epub 2015 Jan 29. Mol Plant Pathol. 2015. PMID: 25469869 Free PMC article. Review.
Cited by
-
Dimerization of the Pseudomonas aeruginosa translocator chaperone PcrH is required for stability, not function.J Bacteriol. 2013 Nov;195(21):4836-43. doi: 10.1128/JB.00335-13. Epub 2013 Aug 23. J Bacteriol. 2013. PMID: 23974025 Free PMC article.
-
Exploring the "N-Terminal Anchor" Binding Interface of the T3SS Chaperone-Translocator Complexes from P. aeruginosa.Biochemistry. 2023 Apr 18;62(8):1420-1427. doi: 10.1021/acs.biochem.3c00002. Epub 2023 Mar 30. Biochemistry. 2023. PMID: 36996474 Free PMC article.
-
Injection of Pseudomonas aeruginosa Exo toxins into host cells can be modulated by host factors at the level of translocon assembly and/or activity.PLoS One. 2012;7(1):e30488. doi: 10.1371/journal.pone.0030488. Epub 2012 Jan 27. PLoS One. 2012. PMID: 22299042 Free PMC article.
-
Protein export according to schedule: architecture, assembly, and regulation of type III secretion systems from plant- and animal-pathogenic bacteria.Microbiol Mol Biol Rev. 2012 Jun;76(2):262-310. doi: 10.1128/MMBR.05017-11. Microbiol Mol Biol Rev. 2012. PMID: 22688814 Free PMC article. Review.
-
Identification of Vibrio cholerae type III secretion system effector proteins.Infect Immun. 2011 Apr;79(4):1728-40. doi: 10.1128/IAI.01194-10. Epub 2011 Jan 31. Infect Immun. 2011. PMID: 21282418 Free PMC article.
References
-
- Cornelis G. R. (2006) Nat. Rev. Microbiol. 4, 811–825 - PubMed
-
- Marlovits T. C., Kubori T., Lara-Tejero M., Thomas D., Unger V. M., Galán J. E. (2006) Nature 441, 637–640 - PubMed
-
- Blocker A., Jouihri N., Larquet E., Gounon P., Ebel F., Parsot C., Sansonetti P., Allaoui A. (2001) Mol. Microbiol. 39, 652–663 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
