

Solution structure of the dimerization domain of ribosomal protein P2 provides insights for the structural organization of eukaryotic stalk
- PMID: 20385603
- PMCID: PMC2926600
- DOI: 10.1093/nar/gkq231
Solution structure of the dimerization domain of ribosomal protein P2 provides insights for the structural organization of eukaryotic stalk
Abstract
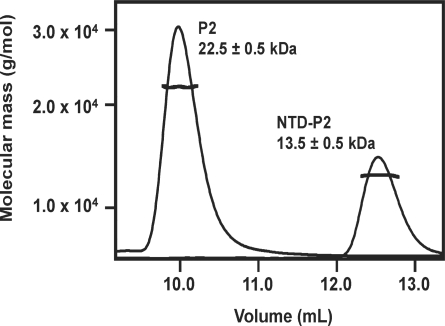
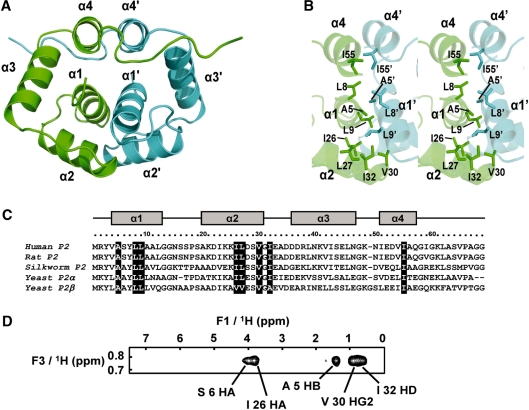
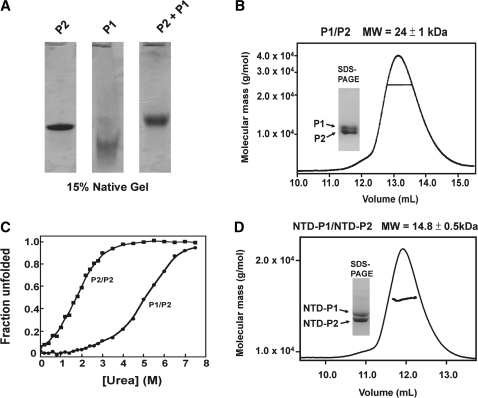
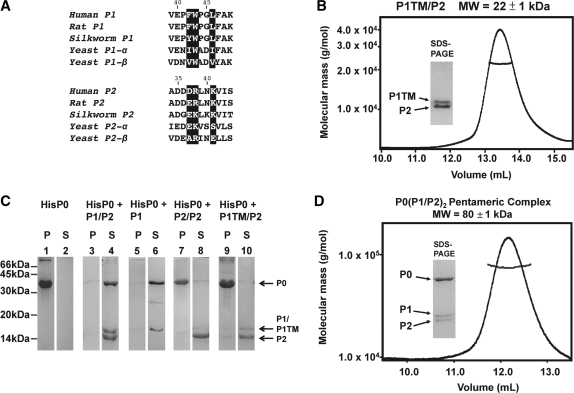
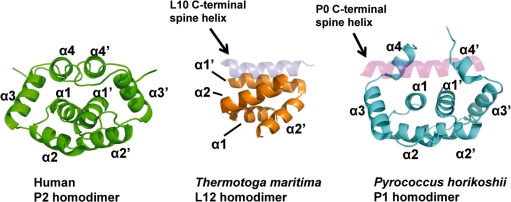
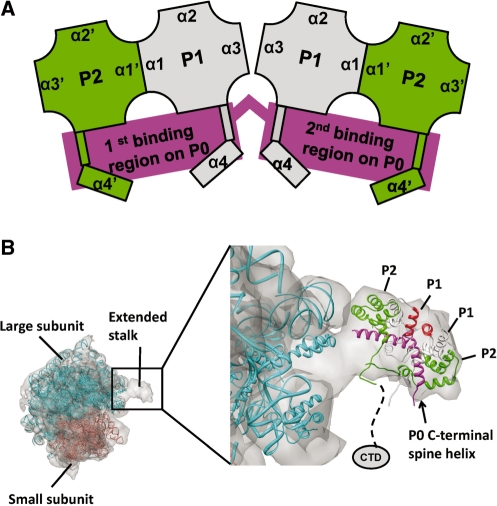
The lateral stalk of ribosome is responsible for kingdom-specific binding of translation factors and activation of GTP hydrolysis that drives protein synthesis. In eukaryotes, the stalk is composed of acidic ribosomal proteins P0, P1 and P2 that constitute a pentameric P-complex in 1: 2: 2 ratio. We have determined the solution structure of the N-terminal dimerization domain of human P2 (NTD-P2), which provides insights into the structural organization of the eukaryotic stalk. Our structure revealed that eukaryotic stalk protein P2 forms a symmetric homodimer in solution, and is structurally distinct from the bacterial counterpart L12 homodimer. The two subunits of NTD-P2 form extensive hydrophobic interactions in the dimeric interface that buries 2400 A(2) of solvent accessible surface area. We have showed that P1 can dissociate P2 homodimer spontaneously to form a more stable P1/P2 1 : 1 heterodimer. By homology modelling, we identified three exposed polar residues on helix-3 of P2 are substituted by conserved hydrophobic residues in P1. Confirmed by mutagenesis, we showed that these residues on helix-3 of P1 are not involved in the dimerization of P1/P2, but instead play a vital role in anchoring P1/P2 heterodimer to P0. Based on our results, models of the eukaryotic stalk complex were proposed.
Figures
Similar articles
-
Solution structure of the dimerization domain of the eukaryotic stalk P1/P2 complex reveals the structural organization of eukaryotic stalk complex.Nucleic Acids Res. 2012 Apr;40(7):3172-82. doi: 10.1093/nar/gkr1143. Epub 2011 Dec 1. Nucleic Acids Res. 2012. PMID: 22135285 Free PMC article.
-
Solution structure of human P1•P2 heterodimer provides insights into the role of eukaryotic stalk in recruiting the ribosome-inactivating protein trichosanthin to the ribosome.Nucleic Acids Res. 2013 Oct;41(18):8776-87. doi: 10.1093/nar/gkt636. Epub 2013 Jul 26. Nucleic Acids Res. 2013. PMID: 23892290 Free PMC article.
-
The amino terminal end determines the stability and assembling capacity of eukaryotic ribosomal stalk proteins P1 and P2.Nucleic Acids Res. 2011 May;39(9):3735-43. doi: 10.1093/nar/gkq1356. Epub 2011 Jan 18. Nucleic Acids Res. 2011. PMID: 21247875 Free PMC article.
-
Proteins P1, P2, and P0, components of the eukaryotic ribosome stalk. New structural and functional aspects.Biochem Cell Biol. 1995 Nov-Dec;73(11-12):959-68. doi: 10.1139/o95-103. Biochem Cell Biol. 1995. PMID: 8722011 Review.
-
Structure and function of the acidic ribosomal stalk proteins.Curr Protein Pept Sci. 2002 Feb;3(1):93-106. doi: 10.2174/1389203023380756. Curr Protein Pept Sci. 2002. PMID: 12370014 Review.
Cited by
-
Crystal Structure of Ribosome-Inactivating Protein Ricin A Chain in Complex with the C-Terminal Peptide of the Ribosomal Stalk Protein P2.Toxins (Basel). 2016 Oct 13;8(10):296. doi: 10.3390/toxins8100296. Toxins (Basel). 2016. PMID: 27754366 Free PMC article.
-
Interaction of ricin and Shiga toxins with ribosomes.Curr Top Microbiol Immunol. 2012;357:1-18. doi: 10.1007/82_2011_174. Curr Top Microbiol Immunol. 2012. PMID: 21910078 Free PMC article. Review.
-
Structural basis of the heterodimerization of the MST and RASSF SARAH domains in the Hippo signalling pathway.Acta Crystallogr D Biol Crystallogr. 2014 Jul;70(Pt 7):1944-53. doi: 10.1107/S139900471400947X. Epub 2014 Jun 29. Acta Crystallogr D Biol Crystallogr. 2014. PMID: 25004971 Free PMC article.
-
Structures of eukaryotic ribosomal stalk proteins and its complex with trichosanthin, and their implications in recruiting ribosome-inactivating proteins to the ribosomes.Toxins (Basel). 2015 Feb 25;7(3):638-47. doi: 10.3390/toxins7030638. Toxins (Basel). 2015. PMID: 25723321 Free PMC article. Review.
-
A Sixty-Year Research and Development of Trichosanthin, a Ribosome-Inactivating Protein.Toxins (Basel). 2022 Feb 27;14(3):178. doi: 10.3390/toxins14030178. Toxins (Basel). 2022. PMID: 35324675 Free PMC article. Review.
References
-
- Uchiumi T, Honma S, Nomura T, Dabbs ER, Hachimori A. Translation elongation by a hybrid ribosome in which proteins at the GTPase center of the Escherichia coli ribosome are replaced with rat counterparts. J. Biol. Chem. 2002;277:3857–3862. - PubMed
-
- Uchiumi T, Hori K, Nomura T, Hachimori A. Replacement of L7/L12.L10 protein complex in Escherichia coli ribosomes with the eukaryotic counterpart changes the specificity of elongation factor binding. J. Biol. Chem. 1999;274:27578–27582. - PubMed
-
- Pettersson I, Hardy SJ, Liljas A. The ribosomal protein L8 is a complex L7/L12 and L10. FEBS Lett. 1976;64:135–138. - PubMed
-
- Diaconu M, Kothe U, Schlunzen F, Fischer N, Harms JM, Tonevitsky AG, Stark H, Rodnina MV, Wahl MC. Structural basis for the function of the ribosomal L7/12 stalk in factor binding and GTPase activation. Cell. 2005;121:991–1004. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
