

Exposure to HIV-1 directly impairs mucosal epithelial barrier integrity allowing microbial translocation
- PMID: 20386714
- PMCID: PMC2851733
- DOI: 10.1371/journal.ppat.1000852
Exposure to HIV-1 directly impairs mucosal epithelial barrier integrity allowing microbial translocation
Abstract
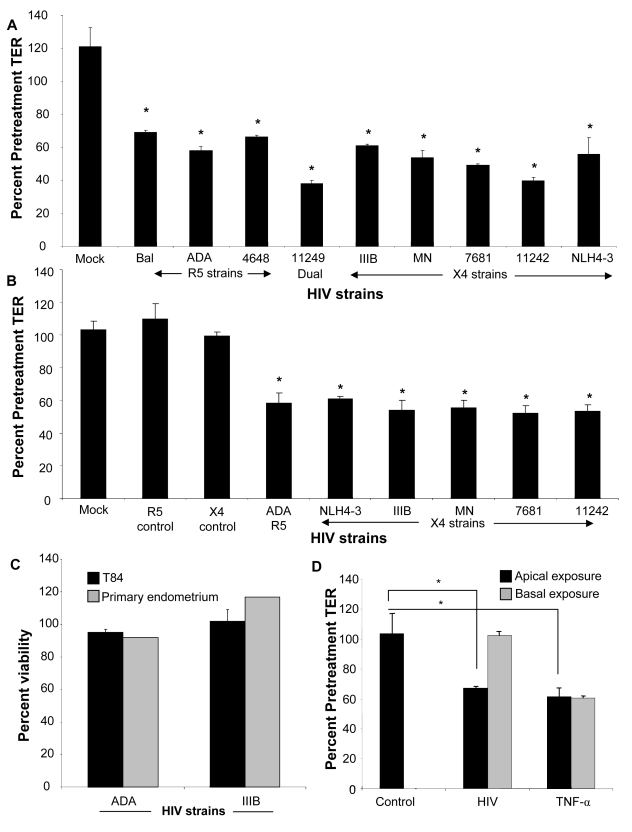
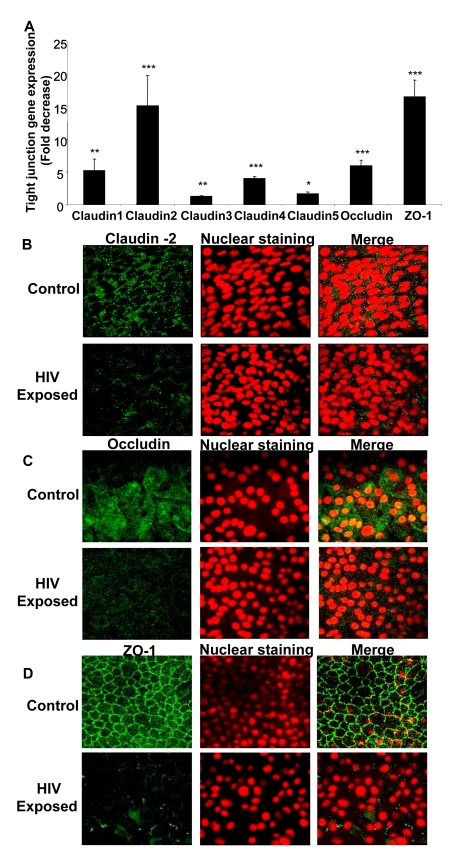
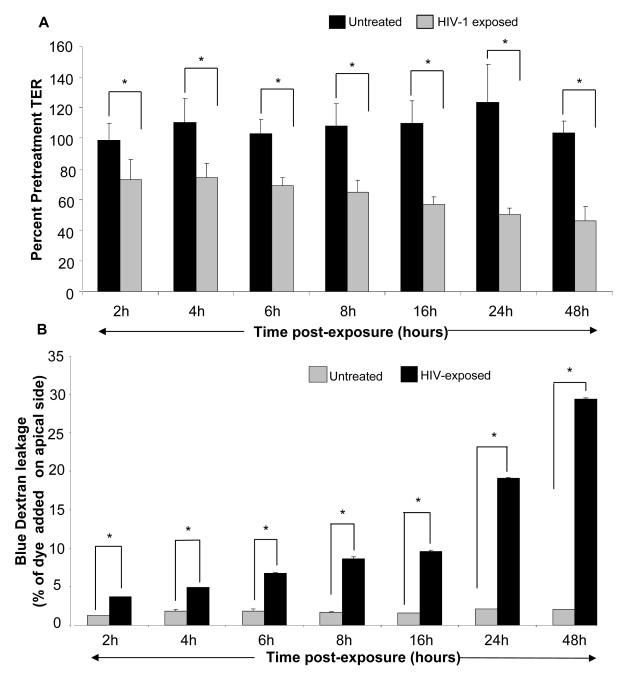
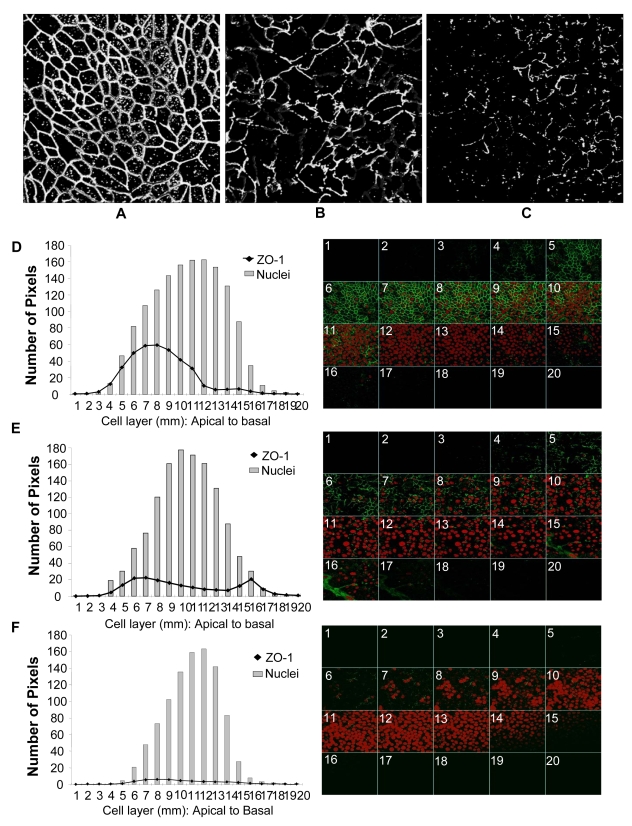
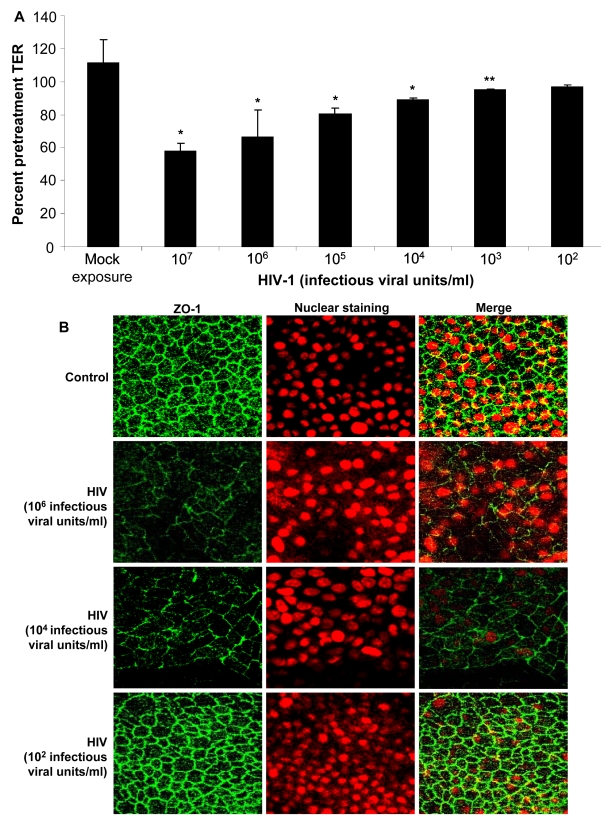
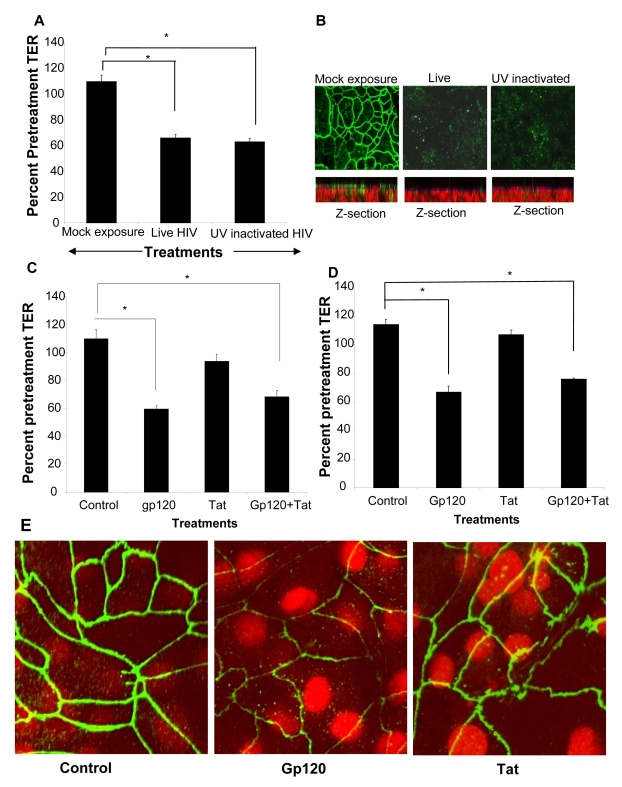
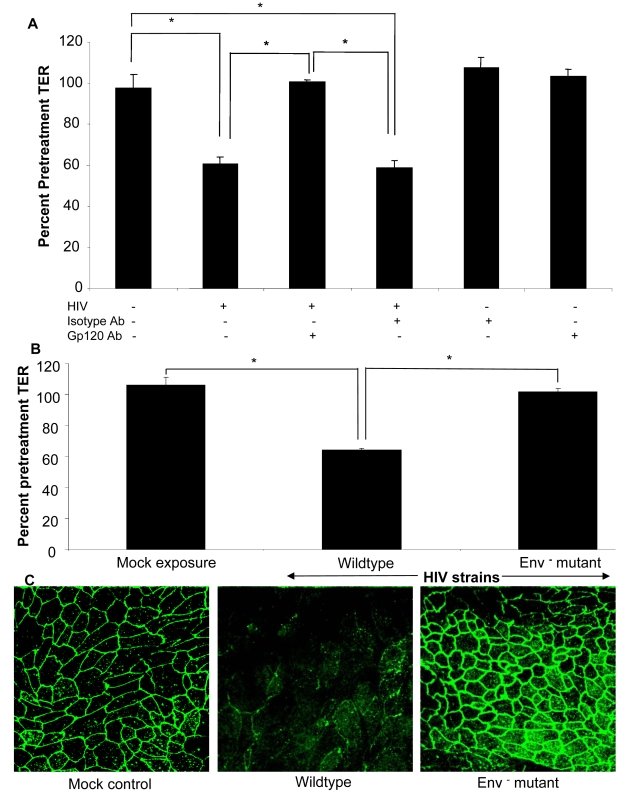
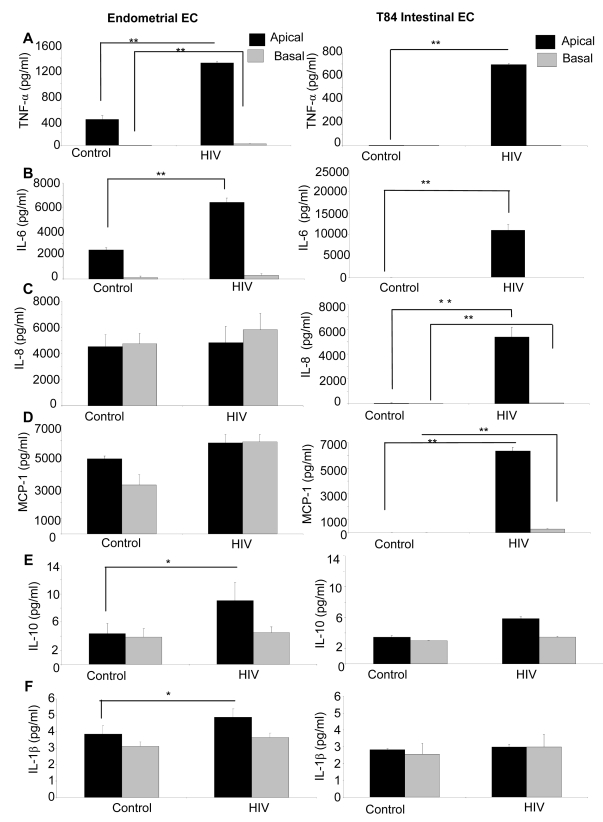
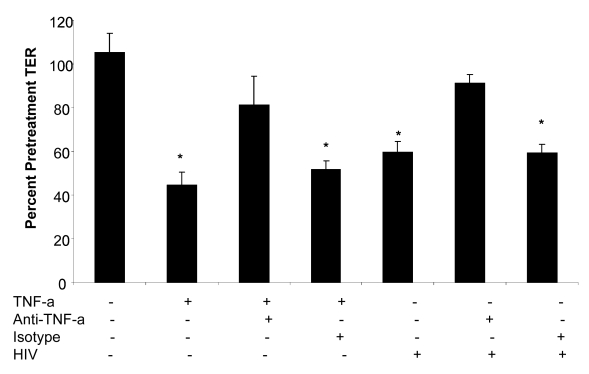
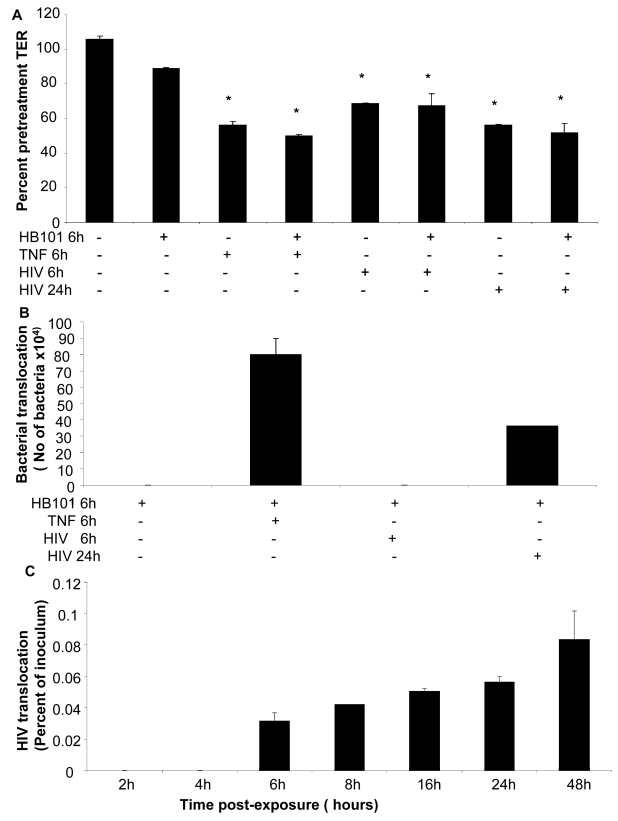
While several clinical studies have shown that HIV-1 infection is associated with increased permeability of the intestinal tract, there is very little understanding of the mechanisms underlying HIV-induced impairment of mucosal barriers. Here we demonstrate that exposure to HIV-1 can directly breach the integrity of mucosal epithelial barrier, allowing translocation of virus and bacteria. Purified primary epithelial cells (EC) isolated from female genital tract and T84 intestinal cell line were grown to form polarized, confluent monolayers and exposed to HIV-1. HIV-1 X4 and R5 tropic laboratory strains and clinical isolates were seen to reduce transepithelial resistance (TER), a measure of monolayer integrity, by 30-60% following exposure for 24 hours, without affecting viability of cells. The decrease in TER correlated with disruption of tight junction proteins (claudin 1, 2, 4, occludin and ZO-1) and increased permeability. Treatment of ECs with HIV envelope protein gp120, but not HIV tat, also resulted in impairment of barrier function. Neutralization of gp120 significantly abrogated the effect of HIV. No changes to the barrier function were observed when ECs were exposed to Env defective mutant of HIV. Significant upregulation of inflammatory cytokines, including TNF-alpha, were seen in both intestinal and genital epithelial cells following exposure to HIV-1. Neutralization of TNF-alpha reversed the reduction in TERs. The disruption in barrier functions was associated with viral and bacterial translocation across the epithelial monolayers. Collectively, our data shows that mucosal epithelial cells respond directly to envelope glycoprotein of HIV-1 by upregulating inflammatory cytokines that lead to impairment of barrier functions. The increased permeability could be responsible for small but significant crossing of mucosal epithelium by virus and bacteria present in the lumen of mucosa. This mechanism could be particularly relevant to mucosal transmission of HIV-1 as well as immune activation seen in HIV-1 infected individuals.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Human immunodeficiency virus-associated disruption of mucosal barriers and its role in HIV transmission and pathogenesis of HIV/AIDS disease.Tissue Barriers. 2016 Mar 3;4(3):e1159276. doi: 10.1080/21688370.2016.1159276. eCollection 2016 Jul-Sep. Tissue Barriers. 2016. PMID: 27583187 Free PMC article. Review.
-
HIV-1 impairs human retinal pigment epithelial barrier function: possible association with the pathogenesis of HIV-associated retinopathy.Lab Invest. 2014 Jul;94(7):777-87. doi: 10.1038/labinvest.2014.72. Epub 2014 May 19. Lab Invest. 2014. PMID: 24840331
-
HIV-1 gp120 induces TLR2- and TLR4-mediated innate immune activation in human female genital epithelium.J Immunol. 2013 Oct 15;191(8):4246-58. doi: 10.4049/jimmunol.1301482. Epub 2013 Sep 16. J Immunol. 2013. PMID: 24043886
-
HIV-1 proteins gp120 and tat induce the epithelial-mesenchymal transition in oral and genital mucosal epithelial cells.PLoS One. 2019 Dec 23;14(12):e0226343. doi: 10.1371/journal.pone.0226343. eCollection 2019. PLoS One. 2019. PMID: 31869348 Free PMC article.
-
Enteric Pathogens and Their Toxin-Induced Disruption of the Intestinal Barrier through Alteration of Tight Junctions in Chickens.Toxins (Basel). 2017 Feb 10;9(2):60. doi: 10.3390/toxins9020060. Toxins (Basel). 2017. PMID: 28208612 Free PMC article. Review.
Cited by
-
Severity of maternal HIV-1 disease is associated with adverse birth outcomes in Malawian women: a cohort study.J Acquir Immune Defic Syndr. 2013 Dec 1;64(4):392-9. doi: 10.1097/QAI.0b013e3182a2d13c. J Acquir Immune Defic Syndr. 2013. PMID: 23846560 Free PMC article.
-
Characterization of a Hormone-Responsive Organotypic Human Vaginal Tissue Model: Morphologic and Immunologic Effects.Reprod Sci. 2015 Aug;22(8):980-90. doi: 10.1177/1933719115570906. Epub 2015 Feb 11. Reprod Sci. 2015. PMID: 25676577 Free PMC article.
-
Isolation and characterization of intestinal epithelial cells from normal and SIV-infected rhesus macaques.PLoS One. 2012;7(1):e30247. doi: 10.1371/journal.pone.0030247. Epub 2012 Jan 26. PLoS One. 2012. PMID: 22291924 Free PMC article.
-
Clinical Assessment and Management in Improving the Quality of Life of HIV/AIDS Patients with Oral Candidiasis: A Case Series.HIV AIDS (Auckl). 2023 Nov 14;15:683-696. doi: 10.2147/HIV.S434175. eCollection 2023. HIV AIDS (Auckl). 2023. PMID: 38028189 Free PMC article.
-
Human immunodeficiency virus-associated disruption of mucosal barriers and its role in HIV transmission and pathogenesis of HIV/AIDS disease.Tissue Barriers. 2016 Mar 3;4(3):e1159276. doi: 10.1080/21688370.2016.1159276. eCollection 2016 Jul-Sep. Tissue Barriers. 2016. PMID: 27583187 Free PMC article. Review.
References
-
- Shattock RJ, Haynes BF, Pulendran B, Flores J, Esparza J. Improving defences at the portal of HIV entry: mucosal and innate immunity. PLoS Med. 2008;5:e81. doi: 10.1371/journal.pmed.0050081. - DOI - PMC - PubMed
-
- Wira CR, Grant-Tschudy KS, Crane-Godreau MA. Epithelial cells in the female reproductive tract: a central role as sentinels of immune protection. Am J Reprod Immunol. 2005;53:65–76. - PubMed
-
- Vroling AB, Fokkens WJ, van Drunen CM. How epithelial cells detect danger: aiding the immune response. Allergy. 2008;63:1110–1123. - PubMed
-
- Yu QH, Yang Q. Diversity of tight junctions (TJs) between gastrointestinal epithelial cells and their function in maintaining the mucosal barrier. Cell Biol Int. 2009;33:78–82. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
