

Cyclic expression of lhx2 regulates hair formation
- PMID: 20386748
- PMCID: PMC2851574
- DOI: 10.1371/journal.pgen.1000904
Cyclic expression of lhx2 regulates hair formation
Abstract
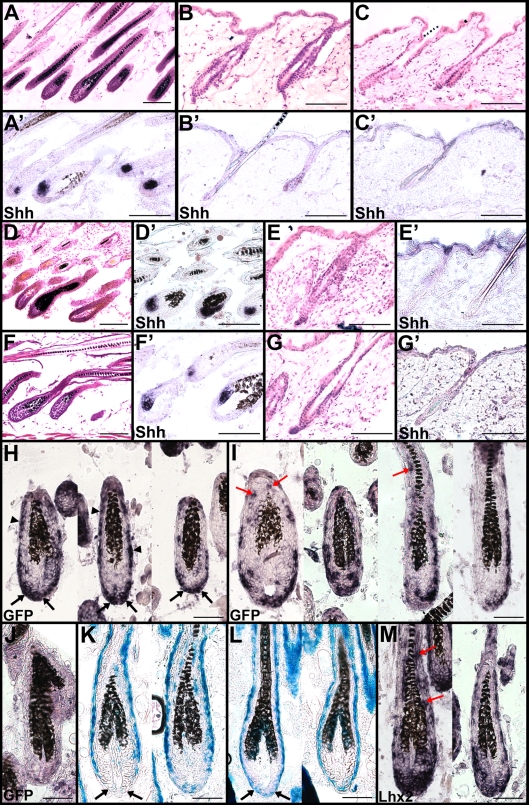
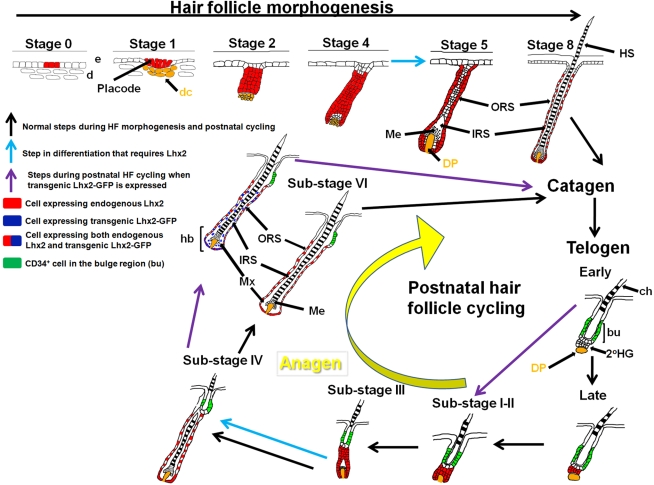
Hair is important for thermoregulation, physical protection, sensory activity, seasonal camouflage, and social interactions. Hair is generated in hair follicles (HFs) and, following morphogenesis, HFs undergo cyclic phases of active growth (anagen), regression (catagen), and inactivity (telogen) throughout life. The transcriptional regulation of this process is not well understood. We show that the transcription factor Lhx2 is expressed in cells of the outer root sheath and a subpopulation of matrix cells during both morphogenesis and anagen. As the HFs enter telogen, expression becomes undetectable and reappears prior to initiation of anagen in the secondary hair germ. In contrast to previously published results, we find that Lhx2 is primarily expressed by precursor cells outside of the bulge region where the HF stem cells are located. This developmental, stage- and cell-specific expression suggests that Lhx2 regulates the generation and regeneration of hair. In support of this hypothesis, we show that Lhx2 is required for anagen progression and HF morphogenesis. Moreover, transgenic expression of Lhx2 in postnatal HFs is sufficient to induce anagen. Thus, our results reveal an alternative interpretation of Lhx2 function in HFs compared to previously published results, since Lhx2 is periodically expressed, primarily in precursor cells distinct from those in the bulge region, and is an essential positive regulator of hair formation.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
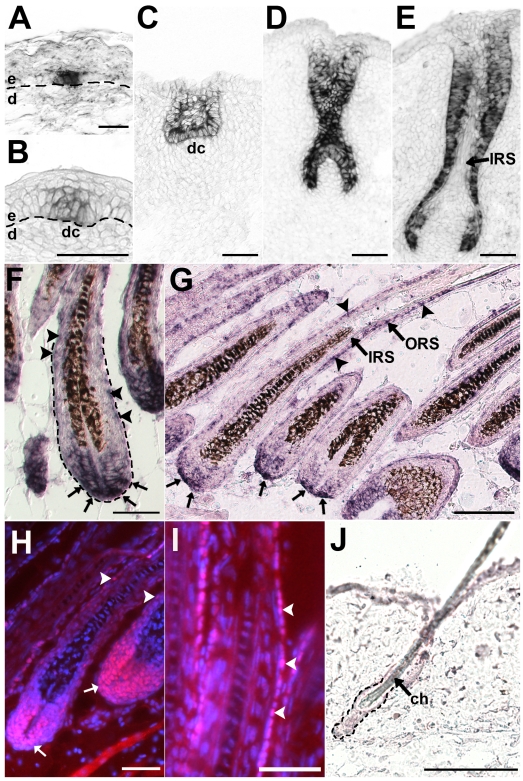
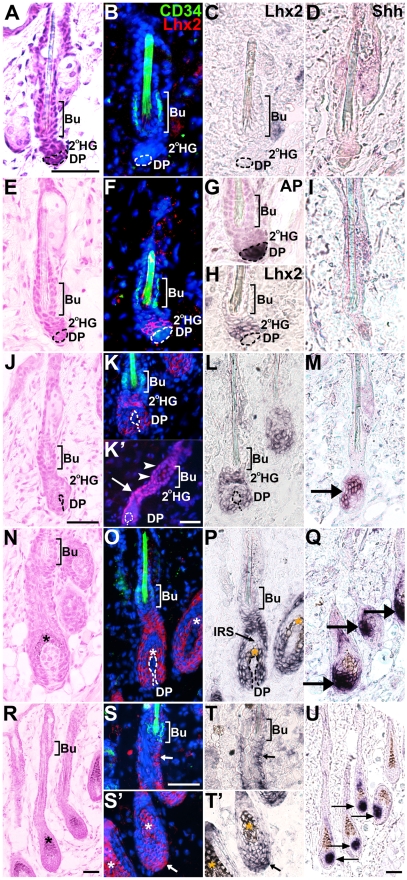
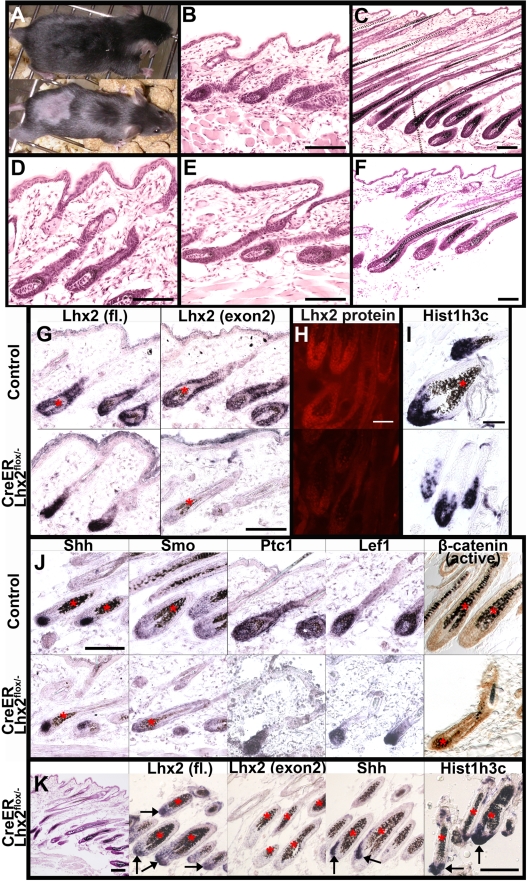
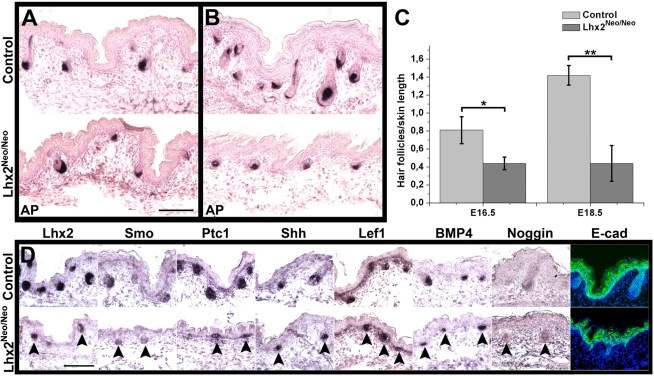
References
-
- Hardy M. The secret life of the hair follicle. Trends Genet. 1992;8:55–61. - PubMed
-
- Botchkarev V, Paus R. Molecular biology of hair morphogenesis: development and cycling. J Exp Zoolog B Mol Dev Evol. 2003;298:164–180. - PubMed
-
- Fuchs E, Merrill B, Jamora C, DasGupta R. At the roots of a never-ending cycle. Develop Cell. 2001;1:13–25. - PubMed
-
- Cotsarelis G, Sun T, Lavker R. Label-retaining cells reside in the bulge area of pilosebaceous unit: implications for follicular stem cells, hair cycle, and skin carcinogenesis. Cell. 1990;61:1329–1337. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
