

Immunomodulatory drugs regulate HMGB1 release from activated human monocytes
- PMID: 20386869
- PMCID: PMC2935946
- DOI: 10.2119/molmed.2010.00031
Immunomodulatory drugs regulate HMGB1 release from activated human monocytes
Abstract
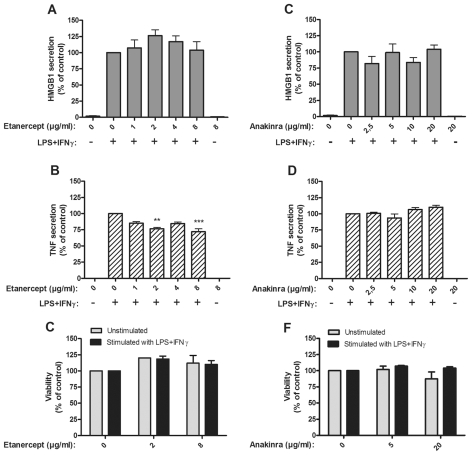
Several HMGB1-specific antagonists have provided beneficial results in multiple models of inflammatory disease-preclinical trials including arthritis. Since no HMGB1-specific targeted therapy has yet reached the clinic, we have performed in vitro studies to investigate whether any of a selection of well-established antirheumatic drugs inhibit HMGB1 release as part of its mode of action. Freshly purified peripheral blood monocytes from healthy donors were stimulated in cultures with LPS and IFNγ to cause HMGB1 and TNF release detected in ELISPOT assays. Effects on the secretion were assessed in cultures supplemented with dexamethasone, cortisone, chloroquine, gold sodium thiomalate, methotrexate, colchicine, etanercept or anakinra. Pharmacologically relevant doses of dexamethasone, gold sodium thiomalate and chloroquine inhibited the extracellular release of HMGB1 in a dose-dependent mode. Immunostaining demonstrated that dexamethasone caused intracellular HMGB1 retention. No effects on HMGB1 secretion were observed in cultures with activated monocytes by any of the other studied agents. TNF production in LPS/IFNγ-activated monocytes was readily downregulated by dexamethasone and, to some extent, by chloroquine and etanercept. We conclude that dexamethasone, gold sodium thiomalate and chloroquine share a capacity to inhibit HMGB1 release from activated monocytes.
Figures
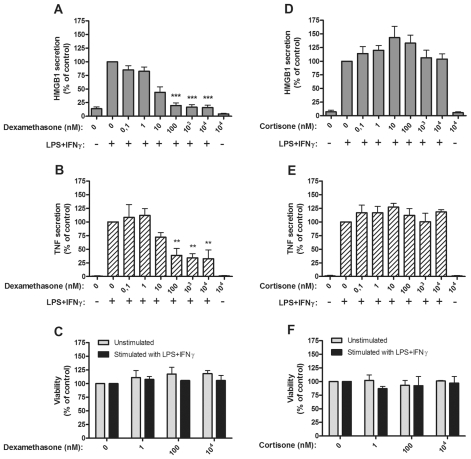
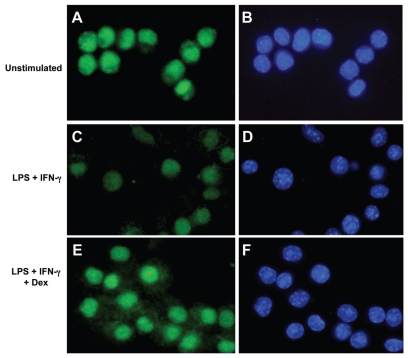
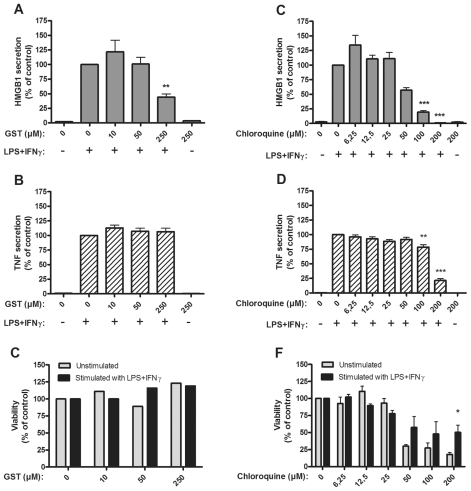
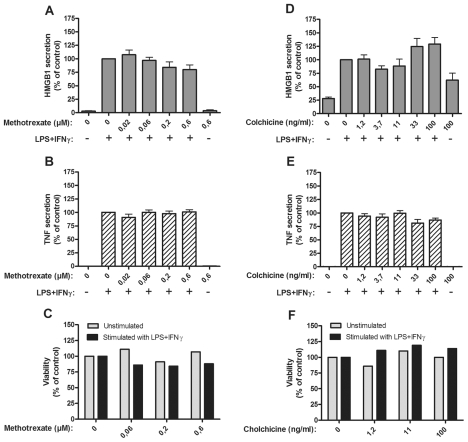
References
-
- Andersson U, Harris HE. The role of HMGB1 in the pathogenesis of rheumatic disease. Biochim. Biophys. Acta. 1799:141–8. - PubMed
-
- Semino C, Angelini G, Poggi A, Rubartelli A. NK/iDC interaction results in IL-18 secretion by DCs at the synaptic cleft followed by NK cell activation and release of the DC maturation factor HMGB1. Blood. 2005;106:609–16. - PubMed
-
- Wang H, et al. HMG-1 as a late mediator of endotoxin lethality in mice. Science. 1999;285:248–51. - PubMed
-
- Wang H, et al. Proinflammatory cytokines (tumor necrosis factor and interleukin 1) stimulate release of high mobility group protein-1 by pituicytes. Surgery. 1999;126:389–92. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
