

Individual carboxypeptidase D domains have both redundant and unique functions in Drosophila development and behavior
- PMID: 20386952
- PMCID: PMC2922403
- DOI: 10.1007/s00018-010-0369-8
Individual carboxypeptidase D domains have both redundant and unique functions in Drosophila development and behavior
Abstract
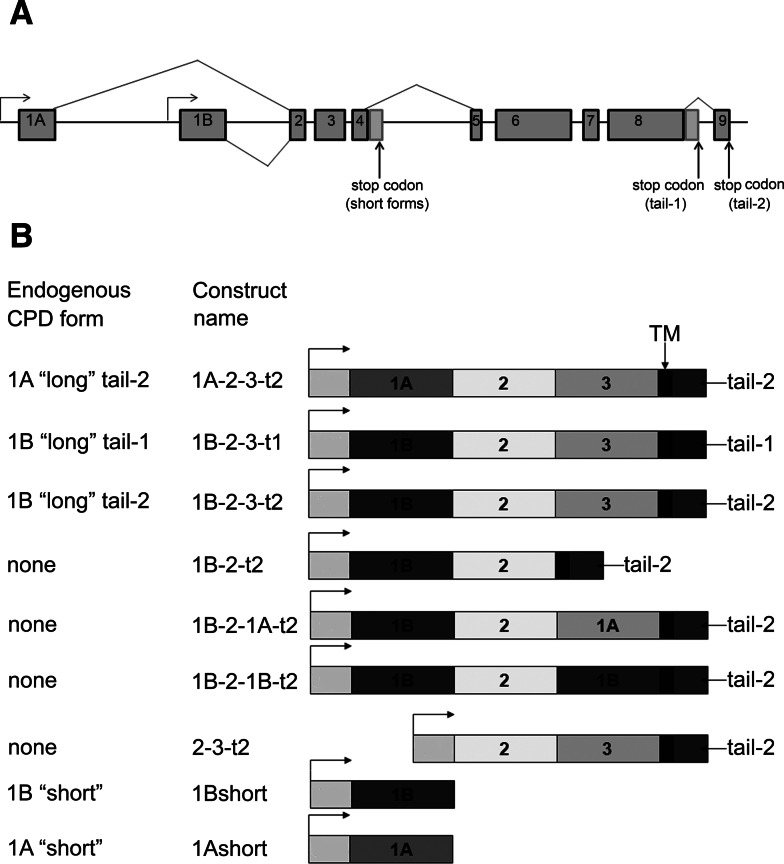
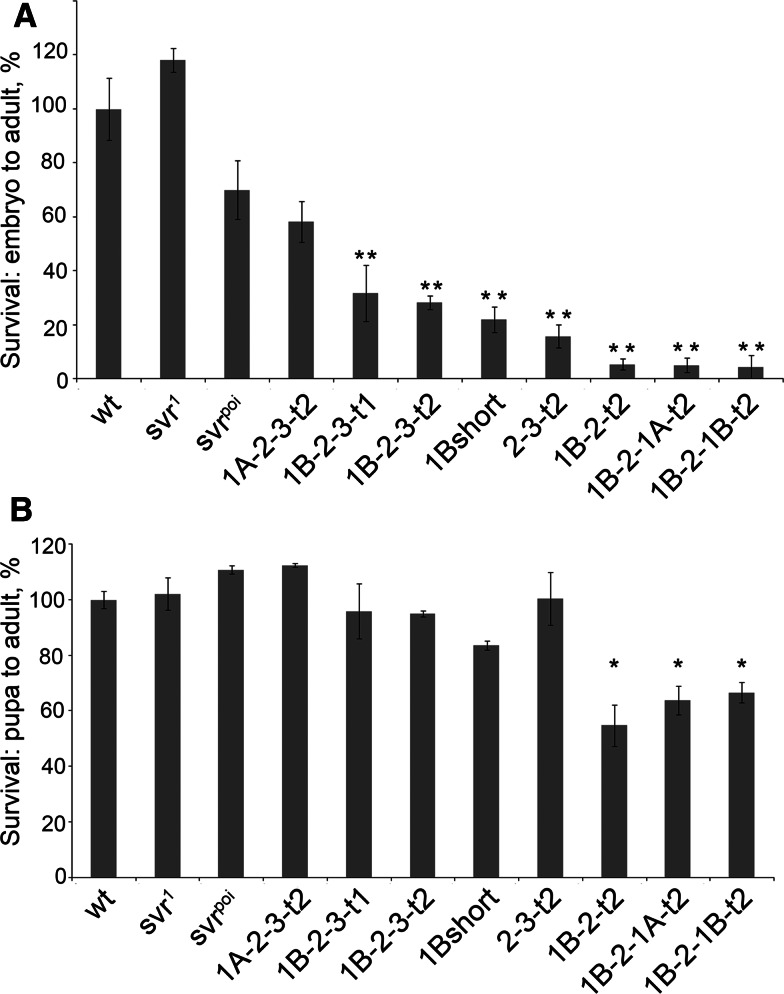
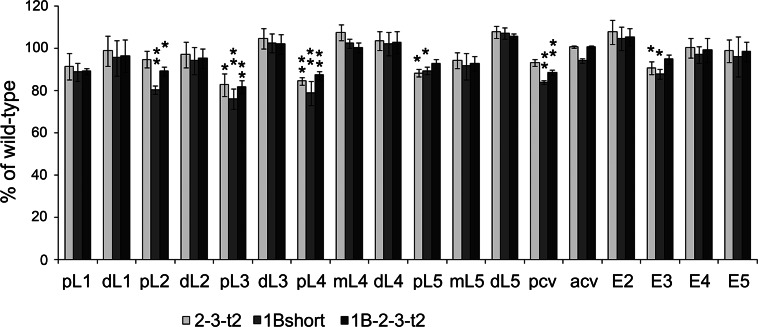
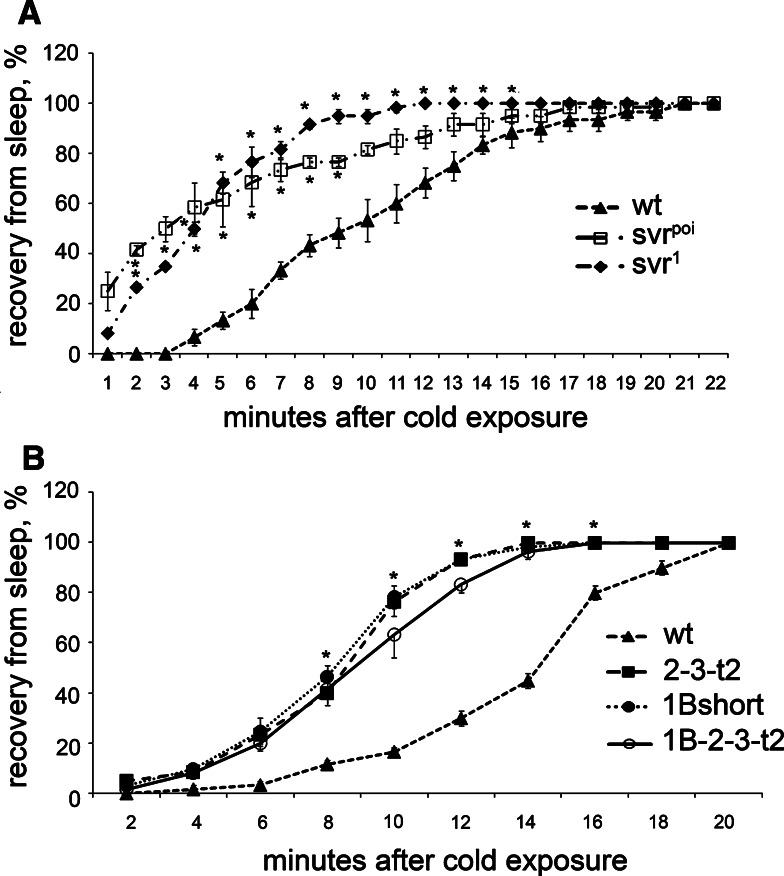
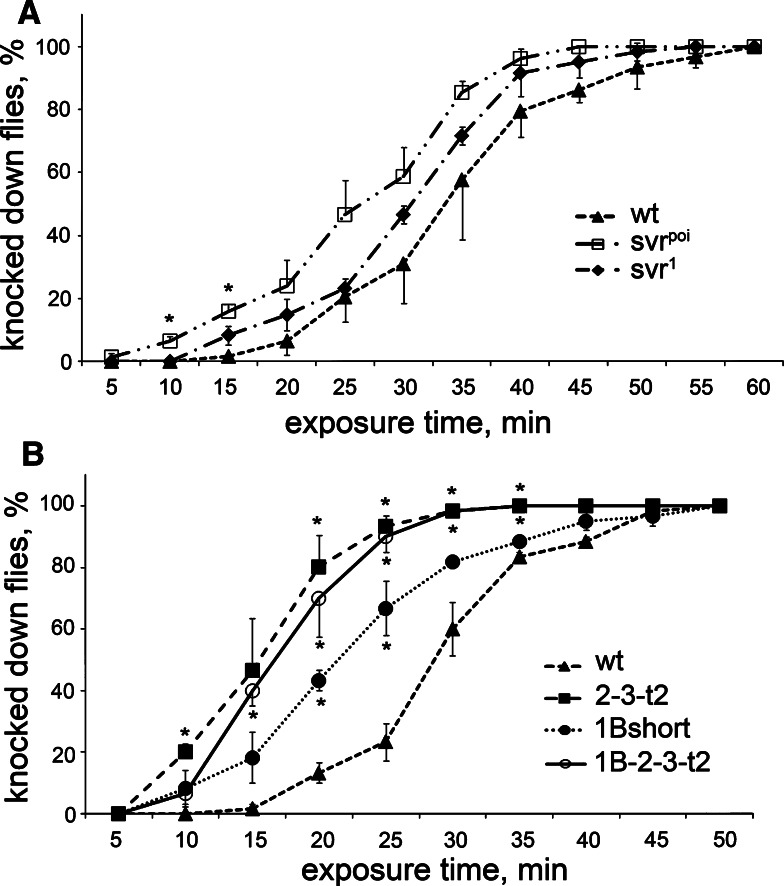
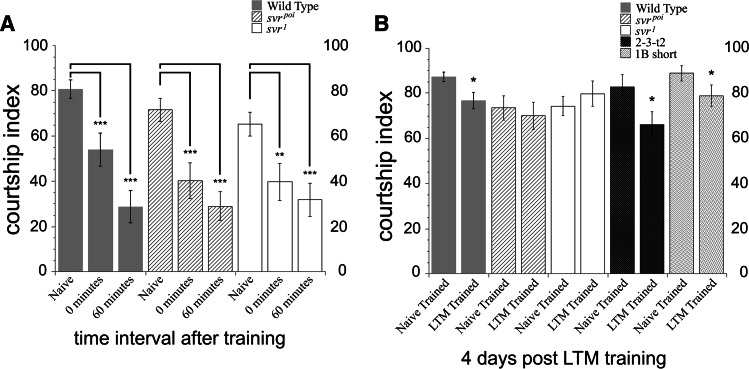
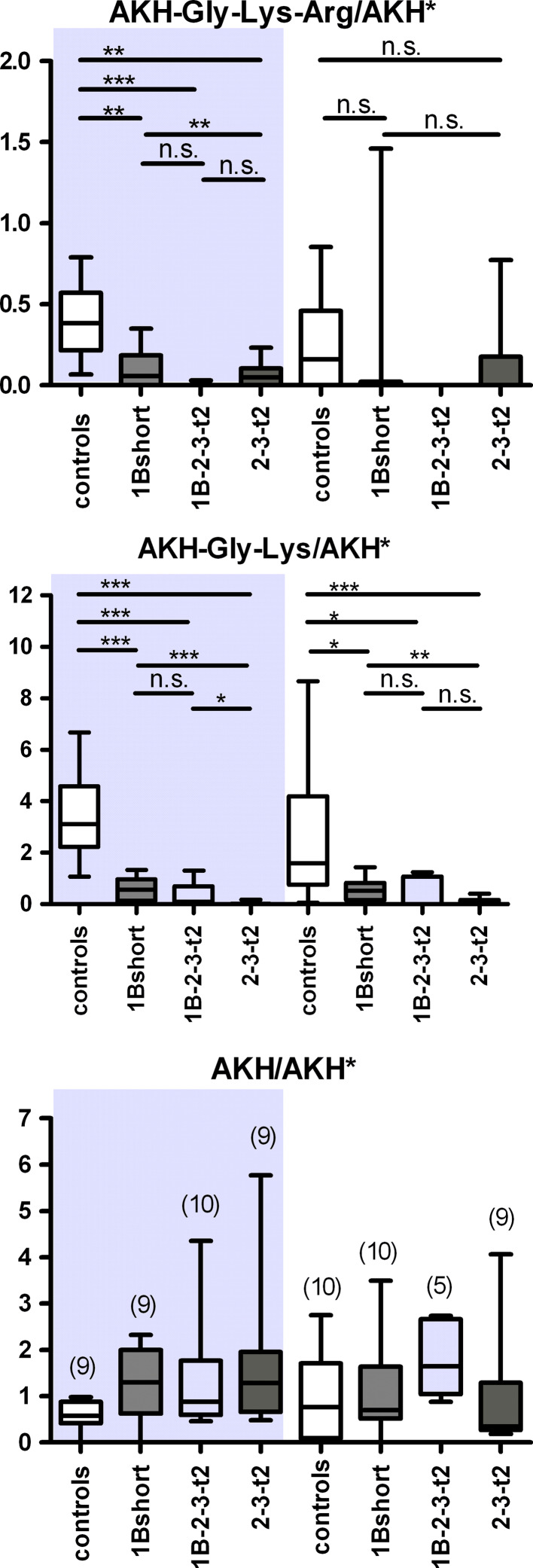
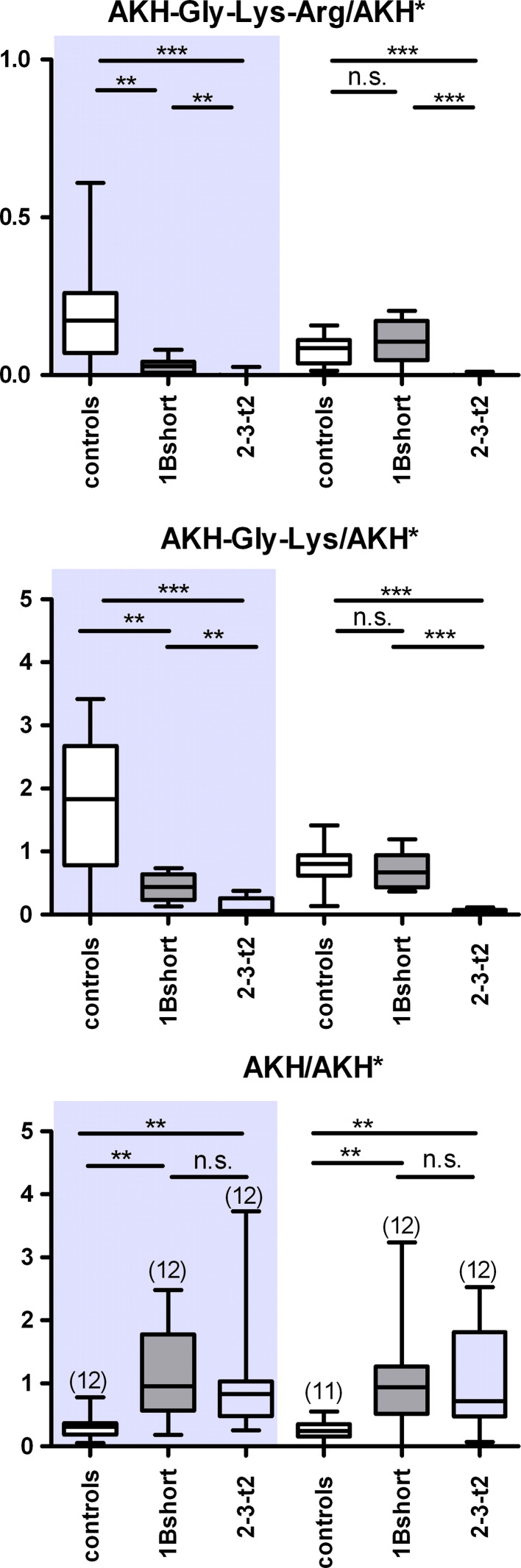
Metallocarboxypeptidase D (CPD) functions in protein and peptide processing. The Drosophila CPD svr gene undergoes alternative splicing, producing forms containing 1-3 active or inactive CP domains. To investigate the function of the various CP domains, we created transgenic flies expressing specific forms of CPD in the embryonic-lethal svr (PG33) mutant. All constructs containing an active CP domain rescued the lethality with varying degrees, and full viability required inactive CP domain-3. Transgenic flies overexpressing active CP domain-1 or -2 were similar to each other and to the viable svr mutants, with pointed wing shape, enhanced ethanol sensitivity, and decreased cold sensitivity. The transgenes fully compensated for a long-term memory deficit observed in the viable svr mutants. Overexpression of CP domain-1 or -2 reduced the levels of Lys/Arg-extended adipokinetic hormone intermediates. These findings suggest that CPD domains-1 and -2 have largely redundant functions in the processing of growth factors, hormones, and neuropeptides.
Figures
References
-
- Strand FL. Neuropeptides: general characteristics and neuropharmaceutical potential in treating CNS disorders. Prog Drug Res. 2003;61:1–37. - PubMed
-
- Rouille Y, Duguay SJ, Lund K, Furuta M, Gong Q, Lipkind G, Oliva AA, Jr, Chan SJ, Steiner DF. Proteolytic processing mechanisms in the biosynthesis of neuroendocrine peptides: the subtilisin-like proprotein convertases. Front Neuroendocrinol. 1995;16:322–361. doi: 10.1006/frne.1995.1012. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
