

Dissociation of calmodulin from cardiac ryanodine receptor causes aberrant Ca(2+) release in heart failure
- PMID: 20388639
- PMCID: PMC3025723
- DOI: 10.1093/cvr/cvq108
Dissociation of calmodulin from cardiac ryanodine receptor causes aberrant Ca(2+) release in heart failure
Abstract
Aims: Calmodulin (CaM) is well known to modulate the channel function of the cardiac ryanodine receptor (RyR2). However, the possible role of CaM on the aberrant Ca(2+) release in diseased hearts remains unclear. In this study, we investigated the state of RyR2-bound CaM and channel dysfunctions in pacing-induced failing hearts.
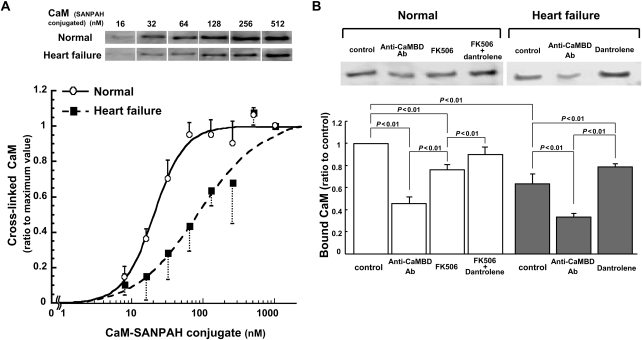
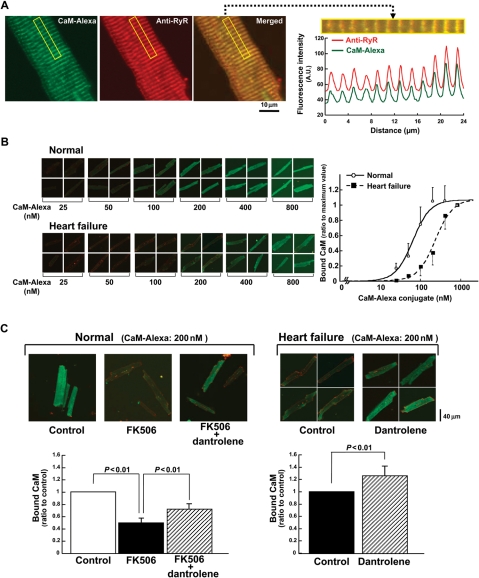
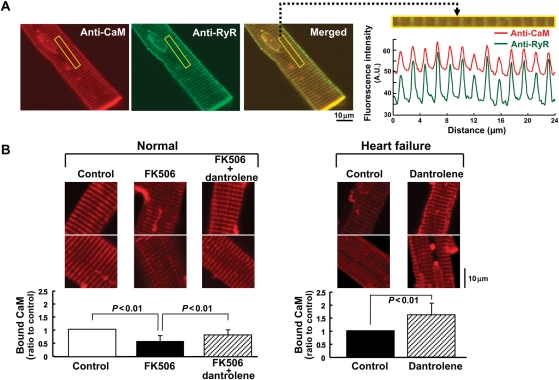
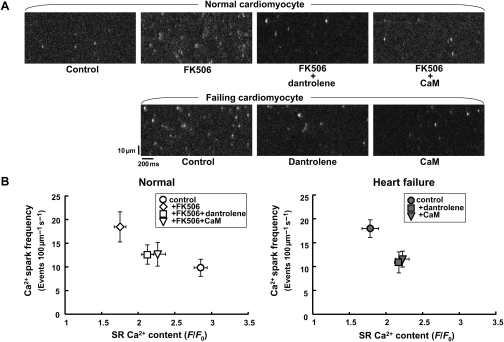
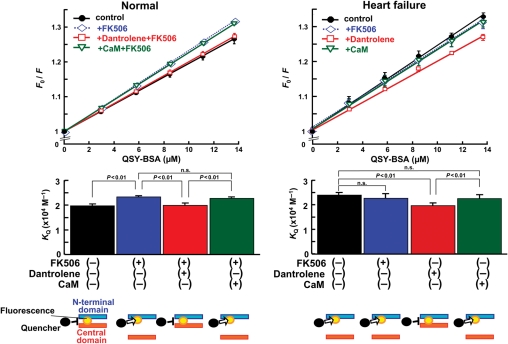
Methods and results: The characteristics of CaM binding to RyR2 and the role of CaM on the aberrant Ca(2+) release were assessed in normal and failing canine hearts. The affinity of CaM binding to RyR2 was lower in failing sarcoplasmic reticulum (SR) than in normal SR. Addition of FK506, which dissociates FKBP12.6 from RyR2, to normal SR reduced the CaM-binding affinity. Dantrolene restored a normal level of the CaM-binding affinity in either FK506-treated (normal) SR or failing SR, suggesting that the defective inter-domain interaction between the N-terminal domain and the central domain of RyR2 (the therapeutic target of dantrolene) is involved in the reduction of the CaM-binding affinity in failing hearts. In saponin-permeabilized cardiomyocytes, the frequency of spontaneous Ca(2+) sparks was much more increased in failing cardiomyocytes than in normal cardiomyocytes, whereas the addition of a high concentration of CaM attenuated the aberrant increase of Ca(2+) sparks.
Conclusion: The defective inter-domain interaction between N-terminal and central domains within RyR2 reduces the binding affinity of CaM to RyR2, thereby causing the spontaneous Ca(2+) release events in failing hearts. Correction of the defective CaM binding may be a new strategy to protect against the aberrant Ca(2+) release in heart failure.
Figures
Comment in
-
Calmodulin: a gatekeeper for ryanodine receptor function in the myocardium.Cardiovasc Res. 2010 Sep 1;87(4):587-8. doi: 10.1093/cvr/cvq215. Epub 2010 Jun 29. Cardiovasc Res. 2010. PMID: 20587508 No abstract available.
Similar articles
-
Enhanced binding of calmodulin to the ryanodine receptor corrects contractile dysfunction in failing hearts.Cardiovasc Res. 2012 Dec 1;96(3):433-43. doi: 10.1093/cvr/cvs271. Epub 2012 Aug 14. Cardiovasc Res. 2012. PMID: 22893680 Free PMC article.
-
Defective domain-domain interactions within the ryanodine receptor as a critical cause of diastolic Ca2+ leak in failing hearts.Cardiovasc Res. 2009 Feb 15;81(3):536-45. doi: 10.1093/cvr/cvn303. Epub 2008 Nov 7. Cardiovasc Res. 2009. PMID: 18996969 Free PMC article.
-
Oxidation of ryanodine receptor (RyR) and calmodulin enhance Ca release and pathologically alter, RyR structure and calmodulin affinity.J Mol Cell Cardiol. 2015 Aug;85:240-8. doi: 10.1016/j.yjmcc.2015.06.009. Epub 2015 Jun 16. J Mol Cell Cardiol. 2015. PMID: 26092277 Free PMC article.
-
The emerging role of calmodulin regulation of RyR2 in controlling heart rhythm, the progression of heart failure and the antiarrhythmic action of dantrolene.Clin Exp Pharmacol Physiol. 2017 Jan;44(1):135-142. doi: 10.1111/1440-1681.12669. Clin Exp Pharmacol Physiol. 2017. PMID: 27626620 Review.
-
Structural instability of ryanodine receptor 2 causes endoplasmic reticulum (ER) dysfunction as well as sarcoplasmic reticulum (SR) dysfunction.J Cardiol. 2025 Jun;85(6):450-457. doi: 10.1016/j.jjcc.2025.02.003. Epub 2025 Feb 8. J Cardiol. 2025. PMID: 39929264 Review.
Cited by
-
CaMKII-dependent phosphorylation of RyR2 promotes targetable pathological RyR2 conformational shift.J Mol Cell Cardiol. 2016 Sep;98:62-72. doi: 10.1016/j.yjmcc.2016.06.007. Epub 2016 Jun 16. J Mol Cell Cardiol. 2016. PMID: 27318036 Free PMC article.
-
Estradiol Treatment Initiated Early After Ovariectomy Regulates Myocardial Gene Expression and Inhibits Diastolic Dysfunction in Female Cynomolgus Monkeys: Potential Roles for Calcium Homeostasis and Extracellular Matrix Remodeling.J Am Heart Assoc. 2018 Nov 6;7(21):e009769. doi: 10.1161/JAHA.118.009769. J Am Heart Assoc. 2018. PMID: 30571375 Free PMC article.
-
EL20, a potent antiarrhythmic compound, selectively inhibits calmodulin-deficient ryanodine receptor type 2.Heart Rhythm. 2018 Apr;15(4):578-586. doi: 10.1016/j.hrthm.2017.12.017. Epub 2017 Dec 14. Heart Rhythm. 2018. PMID: 29248564 Free PMC article.
-
The functional significance of redox-mediated intersubunit cross-linking in regulation of human type 2 ryanodine receptor.Redox Biol. 2020 Oct;37:101729. doi: 10.1016/j.redox.2020.101729. Epub 2020 Sep 15. Redox Biol. 2020. PMID: 32980662 Free PMC article.
-
Cardiac myocyte Z-line calmodulin is mainly RyR2-bound, and reduction is arrhythmogenic and occurs in heart failure.Circ Res. 2014 Jan 17;114(2):295-306. doi: 10.1161/CIRCRESAHA.114.302857. Epub 2013 Nov 1. Circ Res. 2014. PMID: 24186966 Free PMC article.
References
-
- Yano M, Yamamoto T, Ikemoto N, Matsuzaki M. Abnormal ryanodine receptor function in heart failure. Pharmacol Ther. 2005;107:377–391. doi:10.1016/j.pharmthera.2005.04.003. - DOI - PubMed
-
- Marx SO, Reiken S, Hisamatsu Y, Jayaraman T, Burkhoff D, Rosemblit N, et al. PKA phosphorylation dissociates FKBP12.6 from the calcium release channel (ryanodine receptor): defective regulation in failing hearts. Cell. 2000;101:365–376. doi:10.1016/S0092-8674(00)80847-8. - DOI - PubMed
-
- Yano M, Ono K, Ohkusa T, Suetsugu M, Kohno M, Hisaoka T, et al. Altered stoichiometry of FKBP12.6 versus ryanodine receptor as a cause of abnormal Ca2+ leak through ryanodine receptor in heart failure. Circulation. 2000;102:2131–2136. - PubMed
-
- Ai X, Curran JW, Shannon TR, Bers DM, Pogwizd SM. Ca2+/calmodulin-dependent protein kinase modulates cardiac ryanodine receptor phosphorylation and sarcoplasmic reticulum Ca2+ leak in heart failure. Circ Res. 2005;97:1314–1322. doi:10.1161/01.RES.0000194329.41863.89. - DOI - PubMed
-
- Yano M, Yamamoto T, Ikeda Y, Matsuzaki M. Mechanisms of disease: ryanodine receptor defects in heart failure and fatal arrhythmia. Nat Clin Pract Cardiovasc Med. 2006;3:43–52. doi:10.1038/ncpcardio0419. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
