

Review
doi: 10.1104/pp.110.156000.
Epub 2010 Apr 13.
Arabinogalactan-proteins: key regulators at the cell surface?
Affiliations
- PMID: 20388666
- PMCID: PMC2879789
- DOI: 10.1104/pp.110.156000
Item in Clipboard
Review
Arabinogalactan-proteins: key regulators at the cell surface?
Plant Physiol.
2010 Jun.
Erratum in
- Plant Physiol. 2010 Oct;154(2):1012
No abstract available
Figures
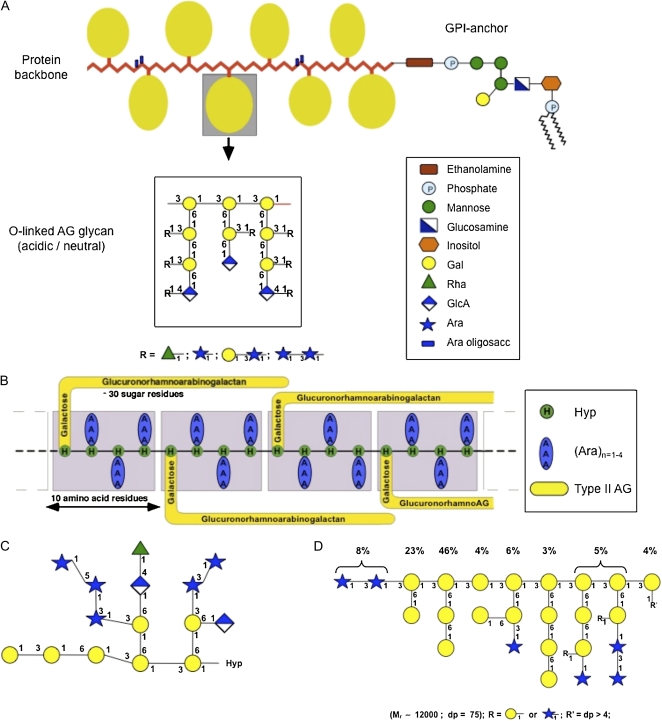
A, The wattle blossom model of the structure of AGPs with a GPI membrane anchor attached. In this model, there are approximately 25 Hyp residues. Most Hyp residues are noncontiguous and are predicted to bear an AG chain. Each AG chain may contain 15 or more repeats of a β-(1-3)-linked Gal oligosaccharide. There may be a few contiguous Hyp residues bearing short arabino-oligosaccharides. The molecule as a whole is spheroidal. The structure of the GPI anchor shows an ethanolamine-phosphate (P) between the anchor and the C terminus of the protein backbone, which is common to all GPI anchors. The core oligosaccharide of the GPI shown is based on PcAGP1 from pear (Pyrus communis; Oxley and Bacic, 1999), which comprises 2- and 6-linked Manp residues, a 4-linked GlcNH2 residue, and a monosubstituted inositol with a partial Galp residue substitution to C(O)4 of the 6-linked Manp residue. The lipid moiety is a ceramide composed primarily of a phytosphingosine base and tetracosanoic acid. This model is modified from Fincher et al. (1983). B, The twisted hairy rope model of the structure of the GAGP. A hypothetical block size of 7 kD contains 10 amino acid residues (1 kD), 30 sugar residues (4.4 kD), and three Hyp-triarabinosides (1.32 kD). The glucuronorhamnoarabinogalactan has a galactan backbone with GlcpA, Rhap, and Araf side chains similar to that shown in the AG schematic in A. This model is from Qi et al. (1991). C, Primary structure of a representative Hyp-AG polysaccharide (AHP-1) released by base hydrolysis from a synthetic AGP (Ala-Hyp)51 from tobacco BY2 cells. This model is modified from Tan et al. (2004). D, Larch AG structure. R′ = dp > 4 is an undefined AG oligosaccharide. This model is modified from Ponder and Richards (1997).
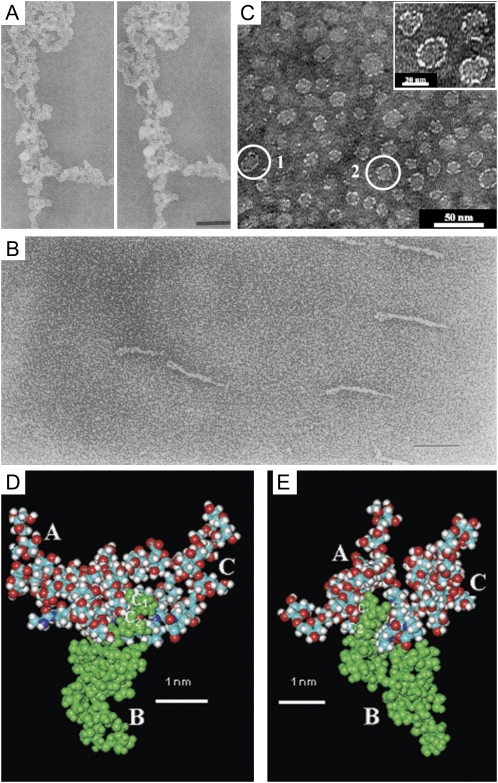
A composite of images of AGPs purified from various plants. A, Stereopair of TEM micrographs showing the three-dimensional structure of a large aggregate of AGP from carrot suspension cultures imaged by the fast-freeze, deep-etch, rotary-shadowed replica technique. Bar = 200 nm. This image is from Baldwin et al. (1993). B, TEM micrograph of Superose-purified GAGP from gum arabic after rotary shadowing. Bar = 100 nm. This image is from Qi et al. (1991). C, TEM micrograph of the AG peptide F1 from acacia gum. The inset shows a magnification of part of the micrograph. This image is modified from Sanchez et al. (2008). D and E, Space-filling CPK models of three AHP-1 AGs (15 glycan residues per polysaccharide chain) from tobacco BY2 cells glycosidically linked to C-4 of each Hyp (O) residue in the 12-residue peptide (A-P-A-O -A-O -A-O -A-P-A-P), in which each O has an AHP-1 substitution (underlined). Nitrogen atoms are shown in dark blue; the oxygen atoms are red; hydrogen atoms are gray; and carbon atoms are turquoise blue. These images are from Tan et al. (2004). D, Glycosylated (A-O/P)6 (see above). Side view of a polysaccharide cluster. Three AHP-1 glycans labeled A to C are O-linked to the Hyp residues of the glycosylated (A-O/P)6 model (residues 4, 6, and 8 of the peptide). The protein backbone lies across the figure, with the N terminus at the far left. Note the close proximity of polysaccharide B (green) to the polypeptide backbone, where the Ara disaccharide residues C1 and C2 form three H bonds as follows: the hydroxymethyl (C-5) of Ara residue C1 to both the carbonyl of Hyp residue 4 and the peptide N of Ala residue 5; and the C-2 hydroxyl of Ara residue C2 to the NH of Ala residue 6. In contrast, the Ara trisaccharide residues at the tip of each polysaccharide form peripheral hook-like projections; these may result in multiple weak interactions (“molecular Velcro”) with the Yariv reagent, which specifically interacts with AGPs. E, Glycopeptide (A-O/P)6. End-on view of a polysaccharide cluster. Reorienting the polypeptide so that it is perpendicular to the plane of the paper shows a syndiotactic propeller-like arrangement of the AG polysaccharides around the polypeptide, providing surfaces for interactions and interdigitation with other matrix molecules.
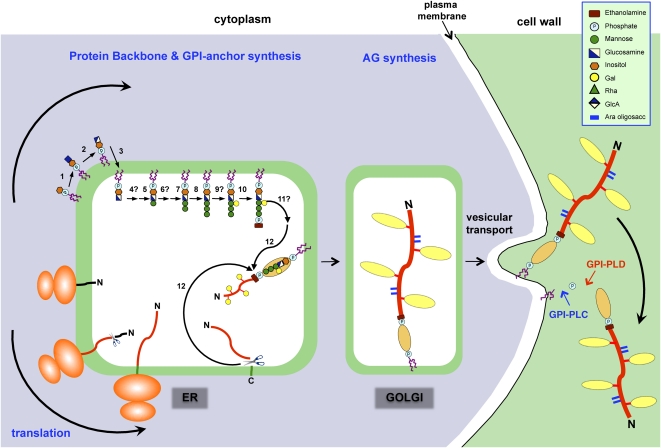
Proposed mechanism for the synthesis and addition of a GPI anchor to an AGP. The addition of a GPI anchor to an AGP likely occurs in several phases, beginning with the synthesis of the GPI moiety on the cytoplasmic surface of the ER. The protein backbone is inserted into the ER cotranslationally, and eventually the two processes converge. The GPI anchor synthesis pathway shown is a composite of that established for mammals and yeast (Orlean and Menon, 2007; Kinoshita et al., 2008) based on the structure of the pear GPI anchor (Fig. 1A). A similar pathway likely exists in plants, since orthologs of the mammalian and yeast genes have been found in plants (Table I). Proteins involved in the biosynthesis pathway (numbered) are summarized in Table I. In both mammals and yeast, any branching glycosylation of the trimannosyl core occurs between steps 8 and 10, and although galactosylation is plant specific, it is presumed to occur at the same stage (step 9). Since substitution of inositol (step 6) and the trimannosyl core (step 11) with acyl and ethanolamine phosphate residues, respectively, has not been demonstrated in plant GPIs, these steps are not shown here, although, interestingly, BLAST searches have identified putative acyl transferases (Table I). Following concomitant removal of the C-terminal GPI signal sequence and addition of the GPI anchor (step 12), numerous Pro residues present within the AGP backbone are hydroxylated to Hyp. Substitution of these Hyp residues with AG chains probably begins within the ER and is completed within the Golgi network. AGPs are transported to the plasma membrane via vesicular transport, where they are either temporarily anchored to the plasma membrane before being released by phospholipases (GPI-PLC and GPI-PLD) or endocytosed. This model is modified from Schultz et al. (1998).
References
-
- Andème-Onzighi C, Sivaguru M, Judy-March J, Baskin TI, Driouich A. (2002) The reb1-1 mutation of Arabidopsis alters the morphology of trichoblasts, the expression of arabinogalactan-proteins and the organization of cortical microtubules. Planta 215: 949–958 - PubMed
-
- Andersson-Gunnerås S, Mellerowicz EJ, Love J, Segerman B, Ohmiya Y, Coutinho PM, Nilsson P, Henrissat B, Moritz T, Sundberg B. (2006) Biosynthesis of cellulose-enriched tension wood in Populus: global analysis of transcripts and metabolites identifies biochemical and developmental regulators in secondary wall biosynthesis. Plant J 45: 144–165 - PubMed
-
- Bacic A, Churms SC, Stephen AM, Cohen PB, Fincher GB. (1987) Fine structure of the arabinogalactan-protein from Lolium multiflorum. Carbohydr Res 162: 85–93
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
