

The immunological synapse: a dynamic platform for local signaling
- PMID: 20390326
- PMCID: PMC2874029
- DOI: 10.1007/s10875-010-9393-6
The immunological synapse: a dynamic platform for local signaling
Abstract
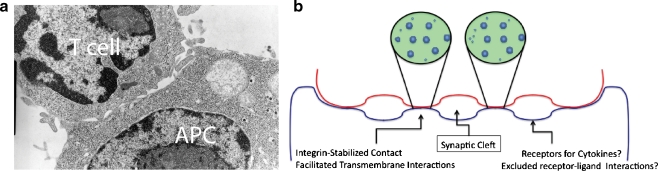
The immunological synapse (IS) as a concept has evolved from a static view of the junction between T cells and their antigen-presenting cell partners. The entire process of IS formation and extinction is now known to entail a dynamic reorganization of membrane domains and proteins within and adjacent to those domains.
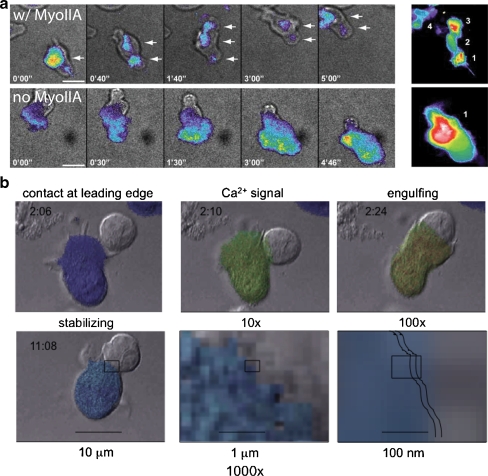
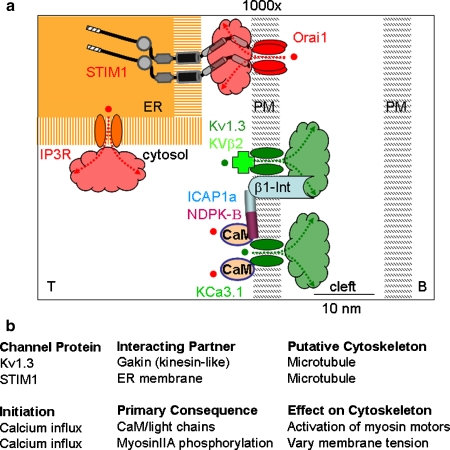
Discussion: The entire process is also intricately tied to the motility machinery-both as that machinery directs "scanning" prior to T-cell receptor engagement and as it is appropriated during the ongoing developments at the IS. While the synapse often remains dynamic in order to encourage surveillance of new antigen-presenting surfaces, cytoskeletal forces also regulate the development of signals, likely including the assembly of ion channels. In both neuronal and immunological synapses, localized Ca2+ signals and accumulation or depletion of ions in microdomains accompany the concentration of signaling molecules in the synapse. Such spatiotemporal signaling in the synapse greatly accelerates kinetics and provides essential checkpoints to validate effective cell-cell communication.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
