

Bone turnover across the menopause transition : The role of gonadal inhibins
- PMID: 20392231
- PMCID: PMC2964283
- DOI: 10.1111/j.1749-6632.2009.05349.x
Bone turnover across the menopause transition : The role of gonadal inhibins
Abstract
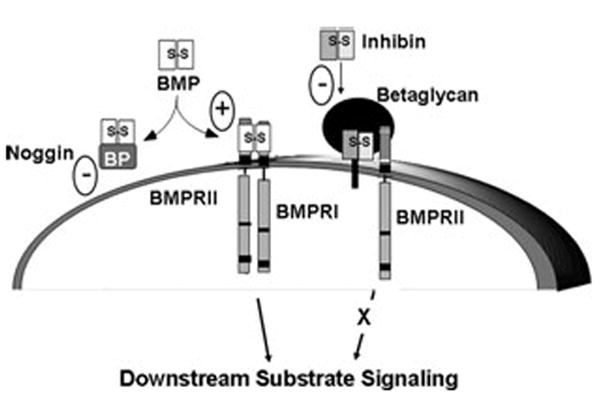
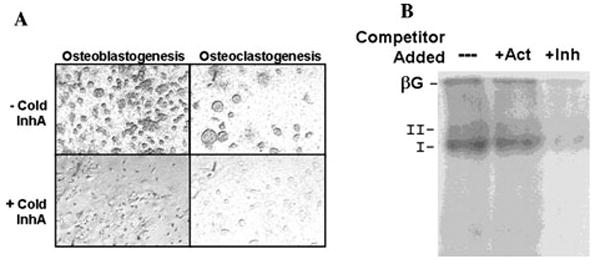
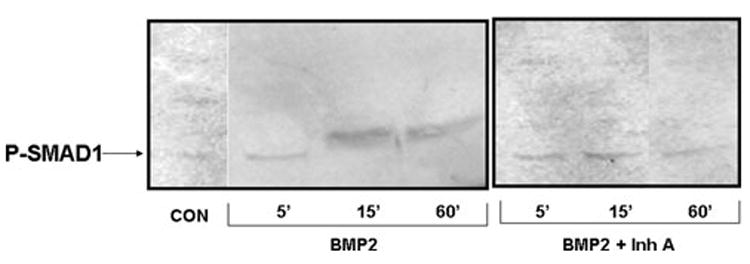
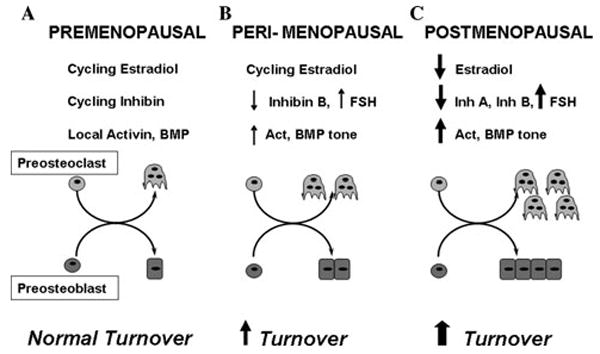
Accumulating evidence demonstrates increasing bone turnover and bone loss in women prior to menopause and decreases in serum estradiol levels. Increased follicle-stimulating hormone levels have been correlated with some of these peri-menopausal changes. However, decreases in gonadal inhibins of the transforming growth factor (TGF)-beta superfamily strongly correlate with increases in bone formation and resorption markers across the menopause transition and predict lumbar bone mass in peri-menopausal women, likely as a result of direct inhibin suppression of osteoblastogenesis and osteoclastogenesis. Inhibins bind specifically to cells during osteoblastogenesis and osteoclastogenesis. They can block bone morphogenetic protein (BMP)-stimulated osteoblast and osteoclast development as well as BMP-stimulated SMAD1 phosphorylation, likely via inhibin-beta-glycan sequestration of BMP Type II receptor (BMPRII). Interestingly, continuous in vivo exposure to inhibin A is anabolic and protective against gonadectomy-induced bone loss in mice, suggesting that inhibins contribute to the endocrine regulation of bone metabolism via a bimodal mechanism of action whereby cycling inhibin exposure suppresses bone turnover and continuous exposure to inhibins is anabolic.
Conflict of interest statement
Figures
References
-
- Albright F, Bloomberg E, Smith PH. Postmenopausal osteoporosis. Trans Assoc Am Phys. 1940;55:298–305.
-
- Manolagas SC. Birth and death of bone cells: basic regulatory mechanisms and implications for the pathogenesis and treatment of osteoporosis. Endocr Rev. 2000;21:115–137. - PubMed
-
- Martin TJ, Sims NA. Osteoclast-derived activity in the coupling of bone formation to resorption. Trends Mol Med. 2005;11:76–81. - PubMed
-
- Manolagas SC, Kousteni S, Jilka RL. Sex steroids and bone. Recent Prog Horm Res. 2002;57:385–409. - PubMed
-
- Riggs BL. Endocrine causes of age-related bone loss and osteoporosis. Novartis Found Symp. 2002;242:247–259. discussion 260–4. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
