

IL-4 directs both CD4 and CD8 T cells to produce Th2 cytokines in vitro, but only CD4 T cells produce these cytokines in response to alum-precipitated protein in vivo
- PMID: 20392496
- PMCID: PMC3826121
- DOI: 10.1016/j.molimm.2010.03.010
IL-4 directs both CD4 and CD8 T cells to produce Th2 cytokines in vitro, but only CD4 T cells produce these cytokines in response to alum-precipitated protein in vivo
Abstract
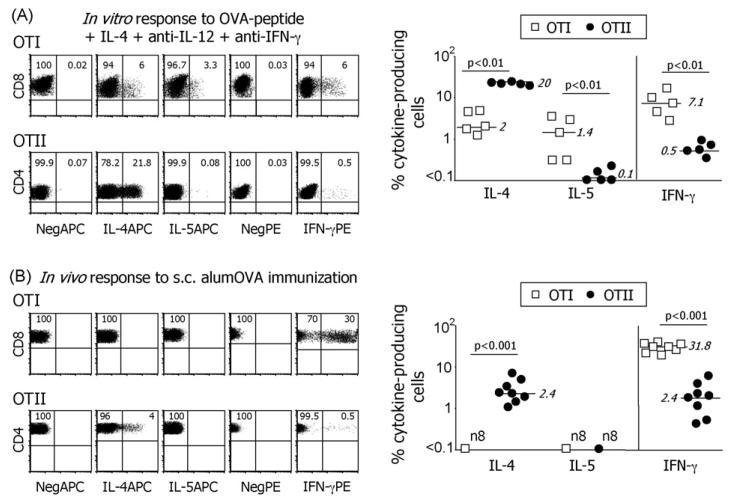
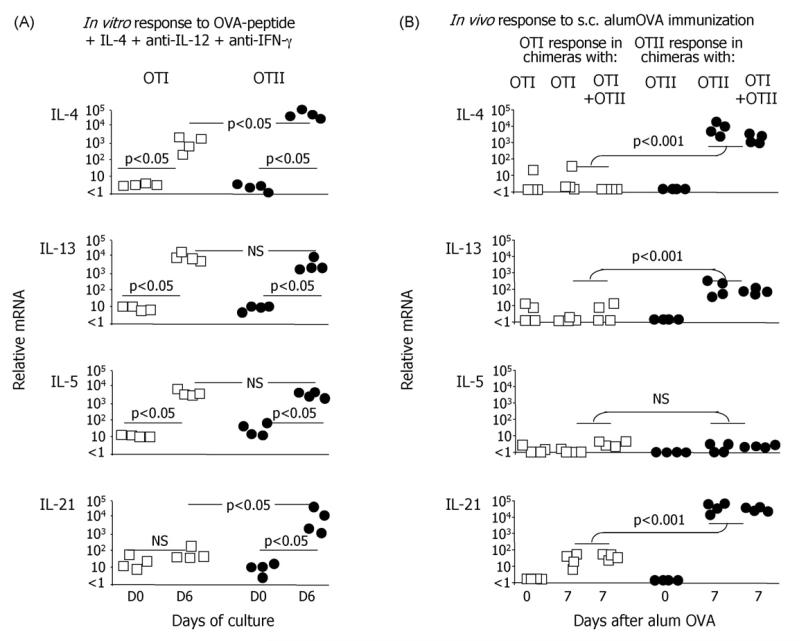
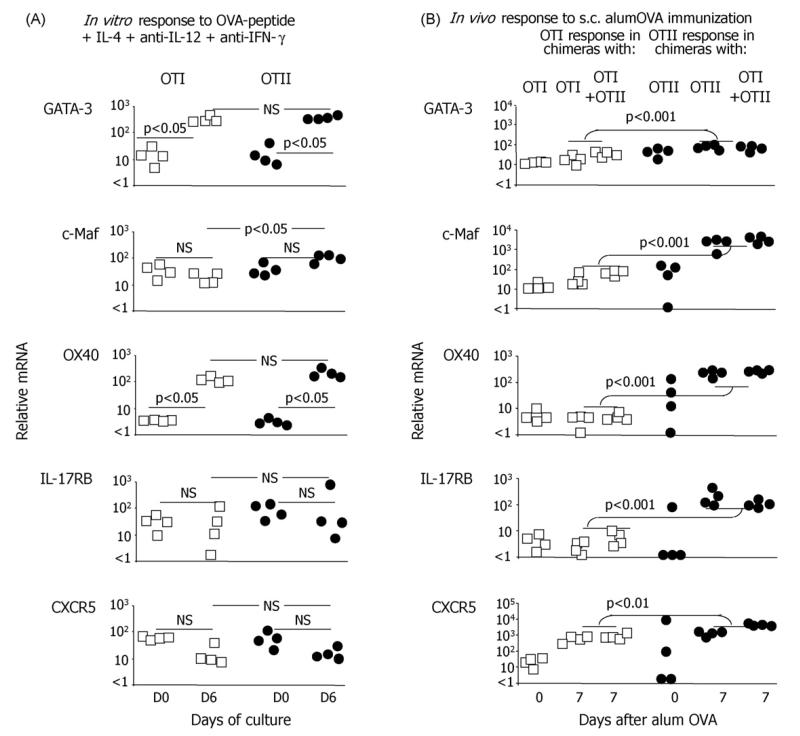
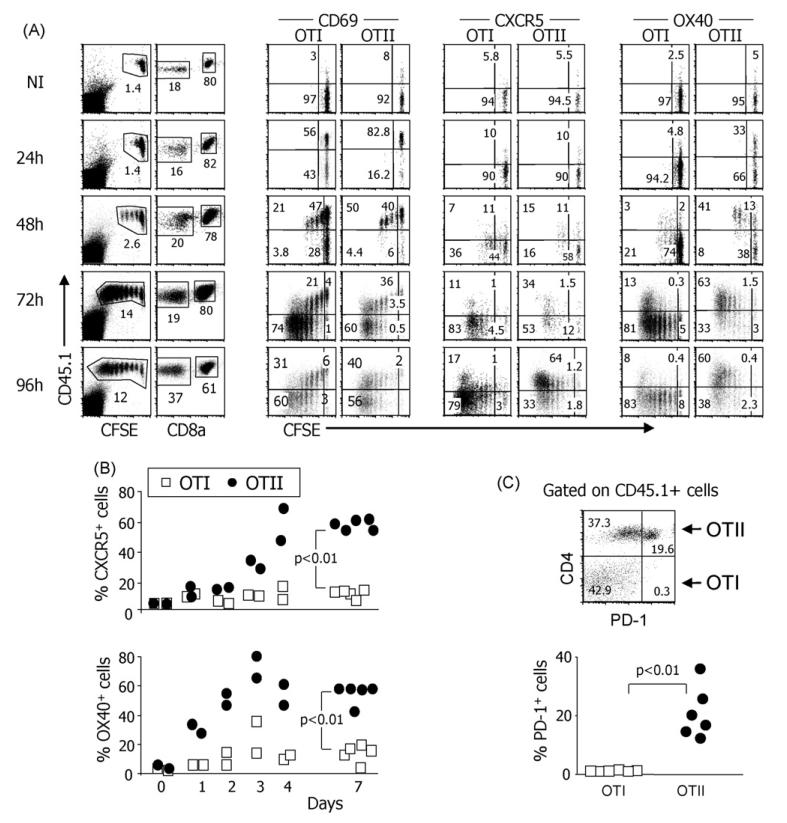
While IL-4 directs CD4 T cells to produce Th2 cytokines (including IL-4, IL-13, IL-5) in vitro it has been shown that production of these cytokines can be induced in vivo in the absence of IL-4/IL-13/STAT-6 signaling. The present report shows that CD8 as well as CD4 T cells activated through their TCR, in vitro upregulate the Th2-features - IL-4, IL-13, IL-5, and GATA-3. However, in vivo while alum-precipitated antigen strongly and selectively induces these Th2-features in CD4 T cells, CD8 T cells mount a markedly different response to this antigen. This CD8 response is associated with strong proliferation and production of IFN-gamma, but no Th2-features are induced. Alum-protein formulations are widely used in human vaccines and typically induce strong antibody responses characterized by the differentiation of IL-4-producing CD4 T cells and immunoglobulin class switching to IgG1. Nevertheless, the mechanism responsible for CD4 Th2 and follicular helper T cell commitment triggered by these alum-protein vaccines is still poorly understood. Analysis of the in vivo response to alum-precipitated protein shows that while subsets of CD4 T cells strongly upregulate Th2 and follicular helper T cell features including the surface markers OX40, CXCR5, PD-1, IL-17RB and the transcription factor c-Maf, CD8 T cells do not. These discrete differences between responding CD4 and CD8 T cells provide further insight into the differences between Th2 polarization of CD4 T cells directed by IL-4 in vitro and the induction of IL-4 production by CD4 T cells in vivo in response to alum-precipitated protein.
Figures
Similar articles
-
Helios is associated with CD4 T cells differentiating to T helper 2 and follicular helper T cells in vivo independently of Foxp3 expression.PLoS One. 2011;6(6):e20731. doi: 10.1371/journal.pone.0020731. Epub 2011 Jun 3. PLoS One. 2011. PMID: 21677778 Free PMC article.
-
Molecular differences between the divergent responses of ovalbumin-specific CD4 T cells to alum-precipitated ovalbumin compared to ovalbumin expressed by Salmonella.Mol Immunol. 2008 Aug;45(13):3558-66. doi: 10.1016/j.molimm.2008.05.010. Epub 2008 Jun 25. Mol Immunol. 2008. PMID: 18582945
-
Selective effects of NF-κB1 deficiency in CD4⁺ T cells on Th2 and TFh induction by alum-precipitated protein vaccines.Eur J Immunol. 2011 Jun;41(6):1573-82. doi: 10.1002/eji.201041126. Epub 2011 May 25. Eur J Immunol. 2011. PMID: 21469117
-
The role of Th2 type CD4+ T cells and Th2 type CD8+ T cells in asthma.Immunol Cell Biol. 1996 Apr;74(2):206-8. doi: 10.1038/icb.1996.29. Immunol Cell Biol. 1996. PMID: 8724011 Review.
-
Inflammatory cytokines as a third signal for T cell activation.Curr Opin Immunol. 2010 Jun;22(3):333-40. doi: 10.1016/j.coi.2010.02.013. Epub 2010 Apr 2. Curr Opin Immunol. 2010. PMID: 20363604 Free PMC article. Review.
Cited by
-
IFN-{gamma} produced by CD8 T cells induces T-bet-dependent and -independent class switching in B cells in responses to alum-precipitated protein vaccine.Proc Natl Acad Sci U S A. 2010 Oct 5;107(40):17292-7. doi: 10.1073/pnas.1004879107. Epub 2010 Sep 20. Proc Natl Acad Sci U S A. 2010. PMID: 20855629 Free PMC article.
-
Mechanisms Underlying the Impact of Interleukin Family on Acute Kidney Injury: Pathogenesis, Progression, and Therapy.Research (Wash D C). 2025 Jun 13;8:0738. doi: 10.34133/research.0738. eCollection 2025. Research (Wash D C). 2025. PMID: 40521043 Free PMC article. Review.
-
Soluble flagellin coimmunization attenuates Th1 priming to Salmonella and clearance by modulating dendritic cell activation and cytokine production.Eur J Immunol. 2015 Aug;45(8):2299-311. doi: 10.1002/eji.201545564. Epub 2015 Jun 24. Eur J Immunol. 2015. PMID: 26036767 Free PMC article.
-
Dendritic cells support a proliferative antigen-specific T-cell response in the presence of poly(lactic-co-glycolic acid).J Biomed Mater Res A. 2021 Nov;109(11):2269-2279. doi: 10.1002/jbm.a.37211. Epub 2021 May 7. J Biomed Mater Res A. 2021. PMID: 33960123 Free PMC article.
-
6-Valent Virus-Like Particle-Based Vaccine Induced Potent and Sustained Immunity Against Noroviruses in Mice.Front Immunol. 2022 May 23;13:906275. doi: 10.3389/fimmu.2022.906275. eCollection 2022. Front Immunol. 2022. PMID: 35711416 Free PMC article.
References
-
- Ballantyne SJ, Barlow JL, Jolin HE, Nath P, Williams AS, Chung KF, Sturton G, Wong SH, McKenzie AN. Blocking IL-25 prevents airway hyperresponsiveness in allergic asthma. J. Allergy Clin. Immunol. 2007;120:1324–1331. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
