

RasGRP1 stimulation enhances ubiquitination and endocytosis of the sodium-chloride cotransporter
- PMID: 20392800
- PMCID: PMC2928521
- DOI: 10.1152/ajprenal.00441.2009
RasGRP1 stimulation enhances ubiquitination and endocytosis of the sodium-chloride cotransporter
Abstract
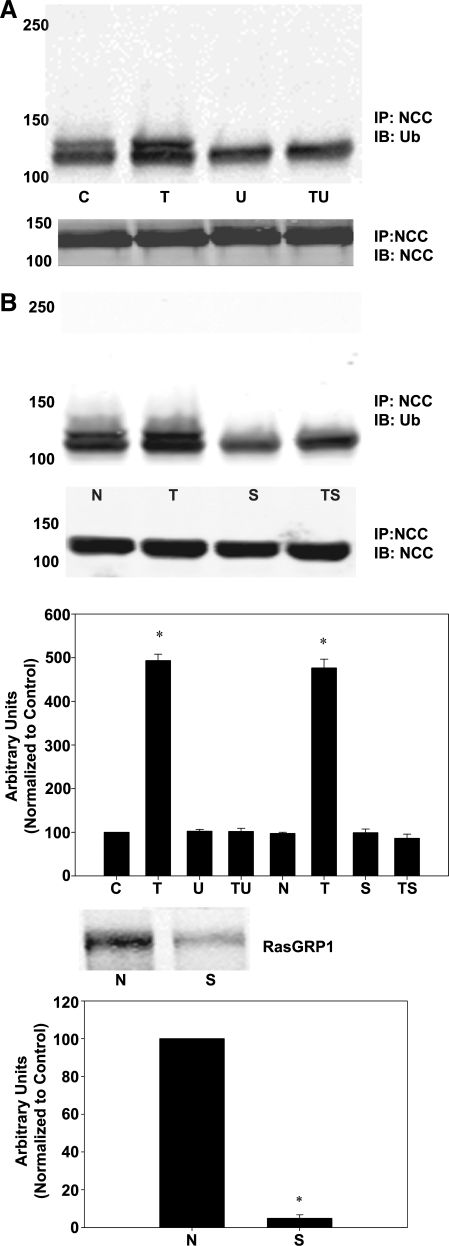
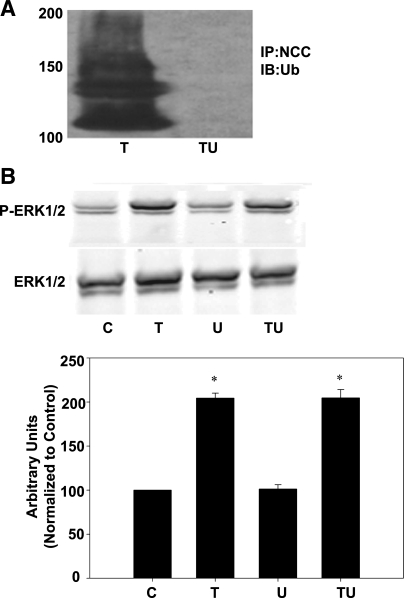
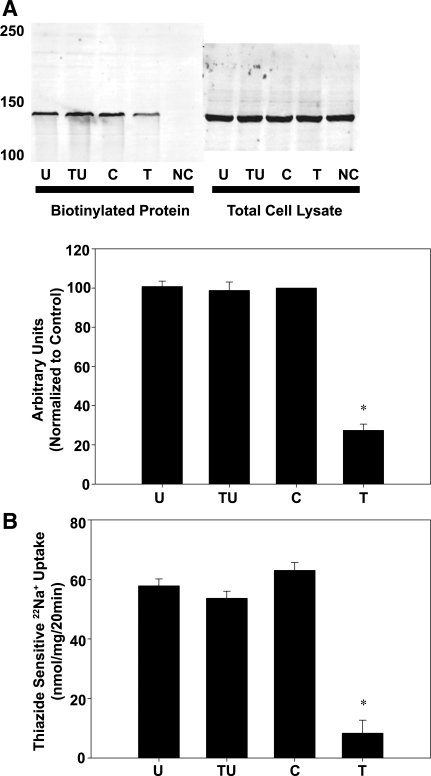
The sodium-chloride cotransporter (NCC) is the principal salt-absorptive pathway in the distal convoluted tubule. Recently, we described a novel pathway of NCC regulation in which phorbol esters (PE) stimulate Ras guanyl-releasing protein 1 (RasGRP1), triggering a cascade ultimately activating ERK1/2 MAPK and decreasing NCC cell surface expression (Ko B, Joshi LM, Cooke LL, Vazquez N, Musch MW, Hebert SC, Gamba G, Hoover RS. Proc Natl Acad Sci USA 104: 20120-20125, 2007). Little is known about the mechanisms which underlie these effects on NCC activity. Regulation of NCC via changes in NCC surface expression has been reported, but endocytosis of NCC has not been demonstrated. In this study, utilizing biotinylation, internalization assays, and a dynamin dominant-negative construct, we demonstrate that the regulation of NCC by PE occurs via an enhancement in internalization of NCC and is dynamin dependent. In addition, immunoprecipitation of NCC and subsequent immunoblotting for ubiquitin showed increased ubiquitination of NCC with phorbol ester treatment. MEK1/2 inhibitors and gene silencing of RasGRP1 indicated that this effect was dependent on RasGRP1 and ERK1/2 activation. Inhibition of ubiquitination prevents any PE-mediated decrease in NCC surface expression as measured by biotinylation or NCC activity as measured by radiotracer uptake. These findings confirmed that the PE effect on NCC is mediated by endocytosis of NCC. Furthermore, ubiquitination of NCC is essential for this process and this ubiquitination is dependent upon RasGRP1-mediated ERK1/2 activation.
Figures
Comment in
-
Regulated endocytosis of NCC.Am J Physiol Renal Physiol. 2010 Aug;299(2):F297-9. doi: 10.1152/ajprenal.00280.2010. Epub 2010 May 26. Am J Physiol Renal Physiol. 2010. PMID: 20504880 Free PMC article. No abstract available.
References
-
- Anonymous Whither RNAi? Nat Cell Biol 5: 489–490, 2003 - PubMed
-
- Booth RE, Stockand JD. Targeted degradation of ENaC in response to PKC activation of the ERK1/2 cascade. Am J Physiol Renal Physiol 284: F938–F947, 2003 - PubMed
-
- Butterworth MB, Edinger RS, Ovaa H, Burg D, Johnson JP, Frizzell RA. The deubiquitinating enzyme UCH-L3 regulates the apical membrane recycling of the epithelial sodium channel. J Biol Chem 282: 37885–37893, 2007 - PubMed
-
- Cai H, Cebotaru V, Wang YH, Zhang XM, Cebotaru L, Guggino SE, Guggino WB. WNK4 kinase regulates surface expression of the human sodium chloride cotransporter in mammalian cells. Kidney Int 69: 2162–2170, 2006 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous