

Structure/Function analysis of the vaccinia virus F18 phosphoprotein, an abundant core component required for virion maturation and infectivity
- PMID: 20392848
- PMCID: PMC2903294
- DOI: 10.1128/JVI.00399-10
Structure/Function analysis of the vaccinia virus F18 phosphoprotein, an abundant core component required for virion maturation and infectivity
Abstract
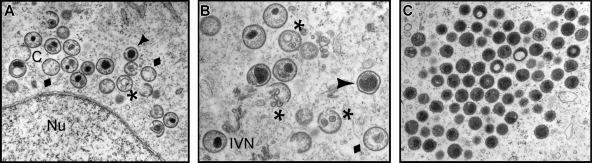
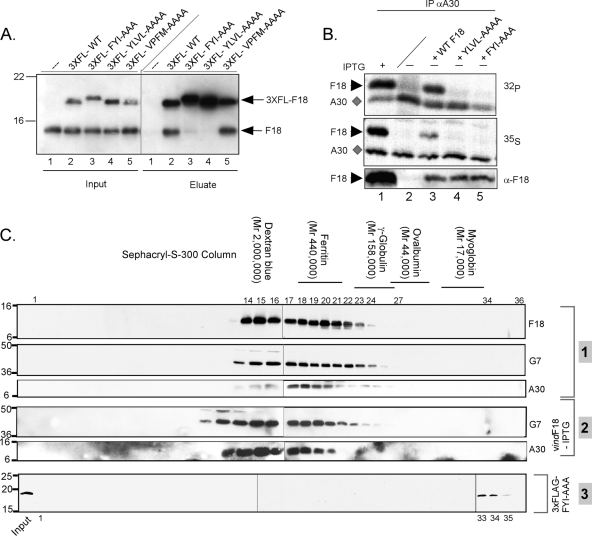
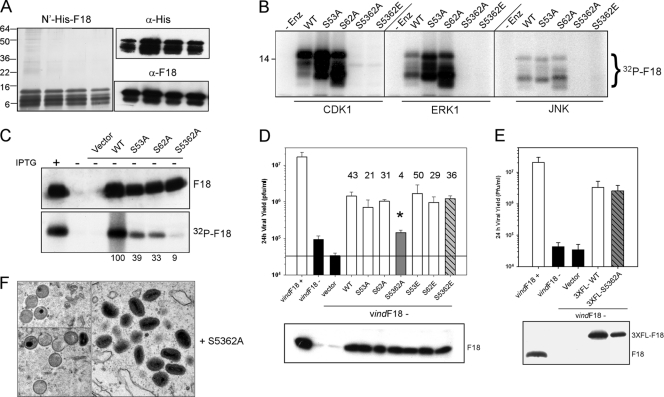
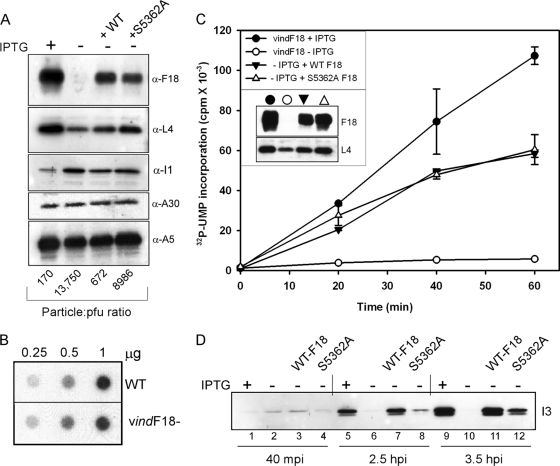
Poxvirus virions, whose outer membrane surrounds two lateral bodies and a core, contain at least 70 different proteins. The F18 phosphoprotein is one of the most abundant core components and is essential for the assembly of mature virions. We report here the results of a structure/function analysis in which the role of conserved cysteine residues, clusters of charged amino acids and clusters of hydrophobic/aromatic amino acids have been assessed. Taking advantage of a recombinant virus in which F18 expression is IPTG (isopropyl-beta-d-thiogalactopyranoside) dependent, we developed a transient complementation assay to evaluate the ability of mutant alleles of F18 to support virion morphogenesis and/or to restore the production of infectious virus. We have also examined protein-protein interactions, comparing the ability of mutant and WT F18 proteins to interact with WT F18 and to interact with the viral A30 protein, another essential core component. We show that F18 associates with an A30-containing multiprotein complex in vivo in a manner that depends upon clusters of hydrophobic/aromatic residues in the N' terminus of the F18 protein but that it is not required for the assembly of this complex. Finally, we confirmed that two PSSP motifs within F18 are the sites of phosphorylation by cellular proline-directed kinases in vitro and in vivo. Mutation of both of these phosphorylation sites has no apparent impact on virion morphogenesis but leads to the assembly of virions with significantly reduced infectivity.
Figures





References
-
- Chakrabarti, S., J. R. Sisler, and B. Moss. 1997. Compact, synthetic, vaccinia virus early/late promoter for protein expression. Biotechniques 23:1094-1097. - PubMed
-
- Condit, R. C., N. Moussatche, and P. Traktman. 2006. In a nutshell: structure and assembly of the vaccinia virion. Adv. Virus Res. 66:31-124. - PubMed
-
- Dyster, L. M., and E. G. Niles. 1991. Genetic and biochemical characterization of vaccinia virus genes D2L and D3R which encode virion structural proteins. Virology 182:455-467. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
