

An antibody directed against the fusion peptide of Junin virus envelope glycoprotein GPC inhibits pH-induced membrane fusion
- PMID: 20392854
- PMCID: PMC2876654
- DOI: 10.1128/JVI.02700-09
An antibody directed against the fusion peptide of Junin virus envelope glycoprotein GPC inhibits pH-induced membrane fusion
Abstract
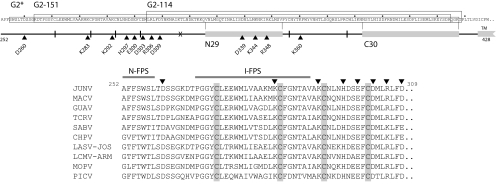
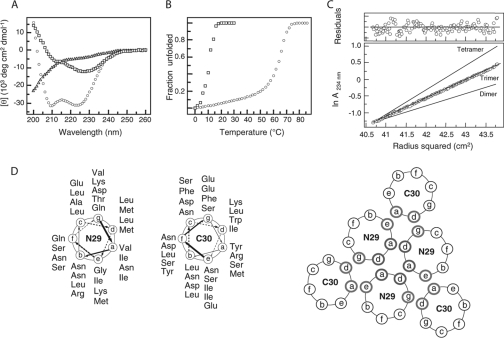
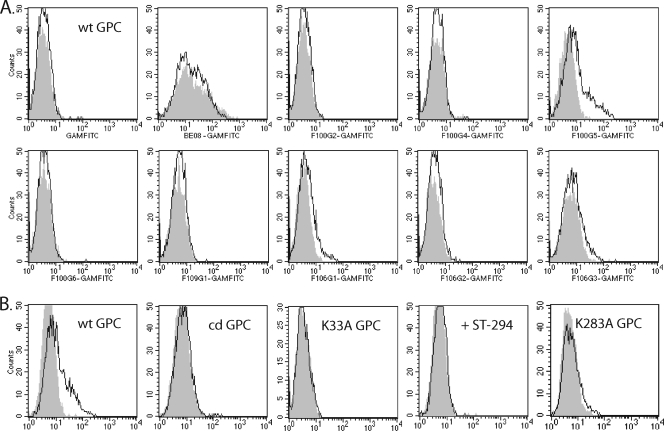
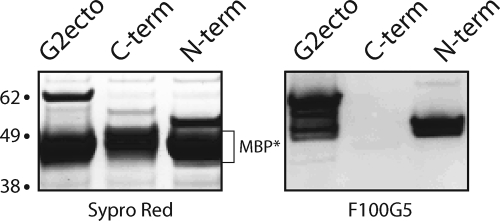
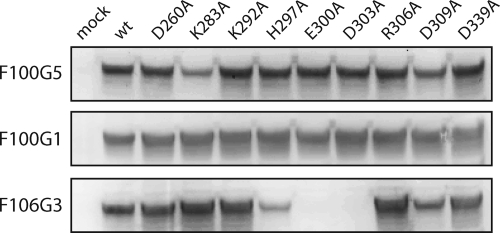
The arenavirus envelope glycoprotein (GPC) initiates infection in the host cell through pH-induced fusion of the viral and endosomal membranes. As in other class I viral fusion proteins, this process proceeds through a structural reorganization in GPC in which the ectodomain of the transmembrane fusion subunit (G2) engages the host cell membrane and subsequently refolds to form a highly stable six-helix bundle structure that brings the two membranes into apposition for fusion. Here, we describe a G2-directed monoclonal antibody, F100G5, that prevents membrane fusion by binding to an intermediate form of the protein on the fusion pathway. Inhibition of syncytium formation requires that F100G5 be present concomitant with exposure of GPC to acidic pH. We show that F100G5 recognizes neither the six-helix bundle nor the larger trimer-of-hairpins structure in the postfusion form of G2. Rather, Western blot analysis using recombinant proteins and a panel of alanine-scanning GPC mutants revealed that F100G5 binding is dependent on an invariant lysine residue (K283) near the N terminus of G2, in the so-called fusion peptide that inserts into the host cell membrane during the fusion process. The F100G5 epitope is located in the internal segment of the bipartite GPC fusion peptide, which also contains four conserved cysteine residues, raising the possibility that this fusion peptide may be highly structured. Collectively, our studies indicate that F100G5 identifies an on-path intermediate form of GPC. Binding to the transiently exposed fusion peptide may interfere with G2 insertion into the host cell membrane. Strategies to effectively target fusion peptide function in the endosome may lead to novel classes of antiviral agents.
Figures
Similar articles
-
Intersubunit interactions modulate pH-induced activation of membrane fusion by the Junin virus envelope glycoprotein GPC.J Virol. 2009 May;83(9):4121-6. doi: 10.1128/JVI.02410-08. Epub 2009 Feb 18. J Virol. 2009. PMID: 19224989 Free PMC article.
-
Myristoylation of the Arenavirus Envelope Glycoprotein Stable Signal Peptide Is Critical for Membrane Fusion but Dispensable for Virion Morphogenesis.J Virol. 2016 Aug 26;90(18):8341-50. doi: 10.1128/JVI.01124-16. Print 2016 Sep 15. J Virol. 2016. PMID: 27412594 Free PMC article.
-
A novel zinc-binding domain is essential for formation of the functional Junín virus envelope glycoprotein complex.J Virol. 2007 Dec;81(24):13385-91. doi: 10.1128/JVI.01785-07. Epub 2007 Oct 10. J Virol. 2007. PMID: 17928348 Free PMC article.
-
The curious case of arenavirus entry, and its inhibition.Viruses. 2012 Jan;4(1):83-101. doi: 10.3390/v4010083. Epub 2012 Jan 13. Viruses. 2012. PMID: 22355453 Free PMC article. Review.
-
Composition and functions of the influenza fusion peptide.Protein Pept Lett. 2009;16(7):766-78. doi: 10.2174/092986609788681715. Protein Pept Lett. 2009. PMID: 19601906 Review.
Cited by
-
Epistastic Interactions within the Junín Virus Envelope Glycoprotein Complex Provide an Evolutionary Barrier to Reversion in the Live-Attenuated Candid#1 Vaccine.J Virol. 2017 Dec 14;92(1):e01682-17. doi: 10.1128/JVI.01682-17. Print 2018 Jan 1. J Virol. 2017. PMID: 29070682 Free PMC article.
-
Biochemical reconstitution of hemorrhagic-fever arenavirus envelope glycoprotein-mediated membrane fusion.PLoS One. 2012;7(11):e51114. doi: 10.1371/journal.pone.0051114. Epub 2012 Nov 30. PLoS One. 2012. PMID: 23226473 Free PMC article.
-
Reduced Susceptibility to VIRIP-Based HIV-1 Entry Inhibitors Has a High Genetic Barrier and Severe Fitness Costs.J Virol. 2018 Aug 16;92(17):e00733-18. doi: 10.1128/JVI.00733-18. Print 2018 Sep 1. J Virol. 2018. PMID: 29925662 Free PMC article.
-
A Cell-Cell Fusion Assay to Assess Arenavirus Envelope Glycoprotein Membrane-Fusion Activity.Methods Mol Biol. 2018;1604:157-167. doi: 10.1007/978-1-4939-6981-4_10. Methods Mol Biol. 2018. PMID: 28986831 Free PMC article.
-
Lymphocytic choriomeningitis virus (LCMV) infection of macaques: a model for Lassa fever.Antiviral Res. 2011 Nov;92(2):125-38. doi: 10.1016/j.antiviral.2011.07.015. Epub 2011 Jul 27. Antiviral Res. 2011. PMID: 21820469 Free PMC article. Review.
References
-
- Barrera Oro, J. G., and K. T. McKee, Jr. 1991. Toward a vaccine against Argentine hemorrhagic fever. Bull. Pan Am. Health Org. 25:118-126. - PubMed
-
- Berry, J. D., S. Jones, M. A. Drebot, A. Andonov, M. Sabara, X. Y. Yuan, H. Weingartl, L. Fernando, P. Marszal, J. Gren, B. Nicolas, M. Andonova, F. Ranada, M. J. Gubbins, T. B. Ball, P. Kitching, Y. Li, A. Kabani, and F. Plummer. 2004. Development and characterisation of neutralising monoclonal antibody to the SARS-coronavirus. J. Virol. Methods 120:87-96. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
