

Combined adaptiveness of specific motor cortical ensembles underlies learning
- PMID: 20392963
- PMCID: PMC6632744
- DOI: 10.1523/JNEUROSCI.0076-10.2010
Combined adaptiveness of specific motor cortical ensembles underlies learning
Abstract
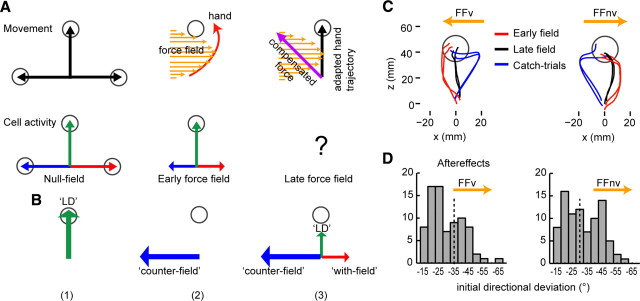
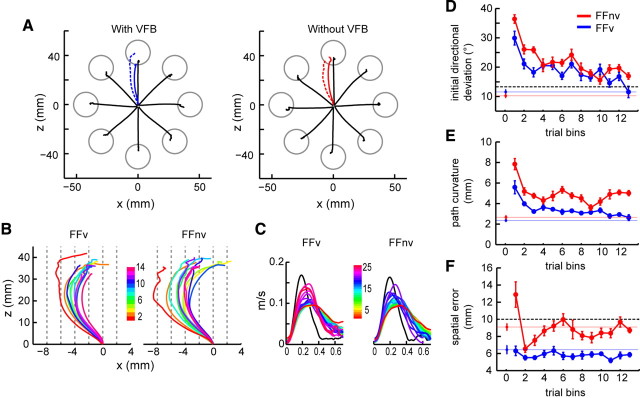
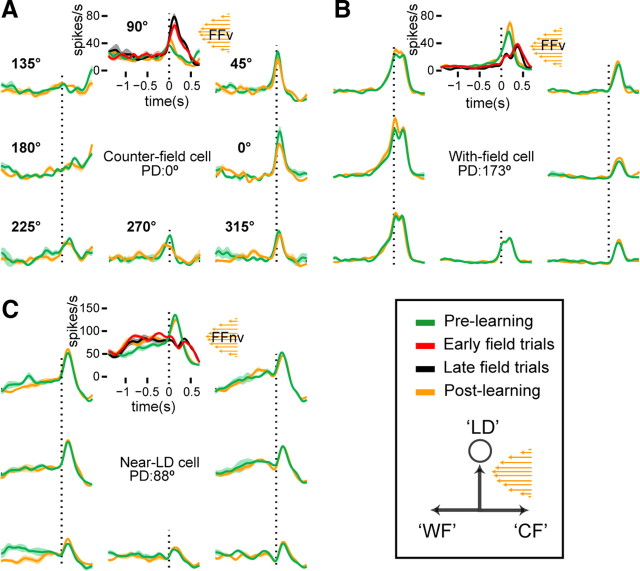
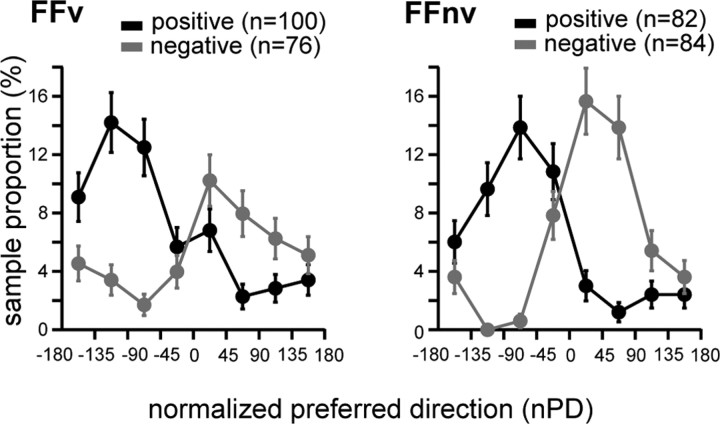
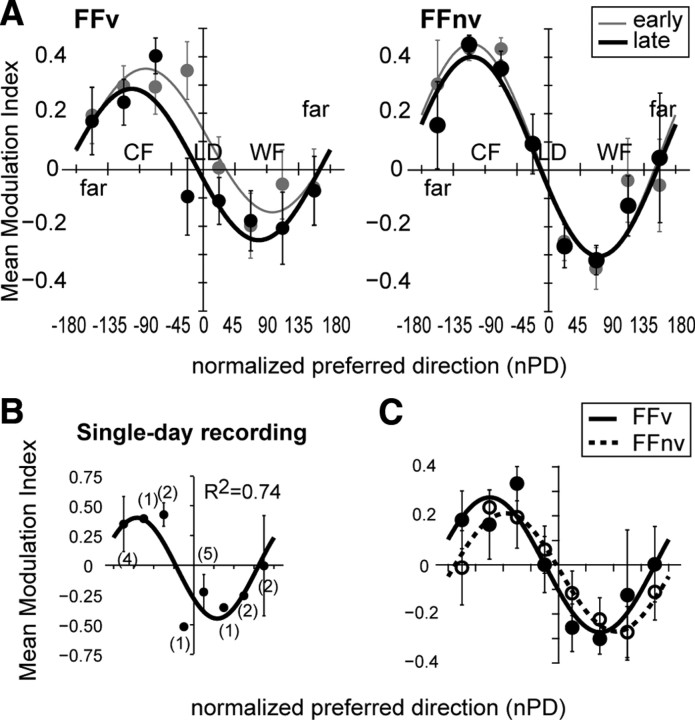
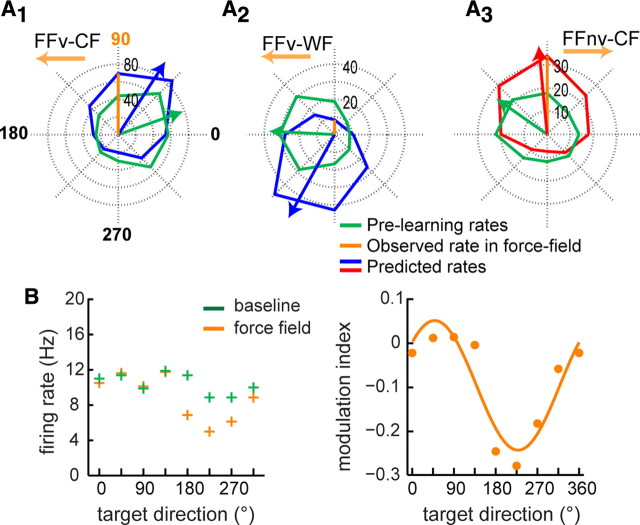
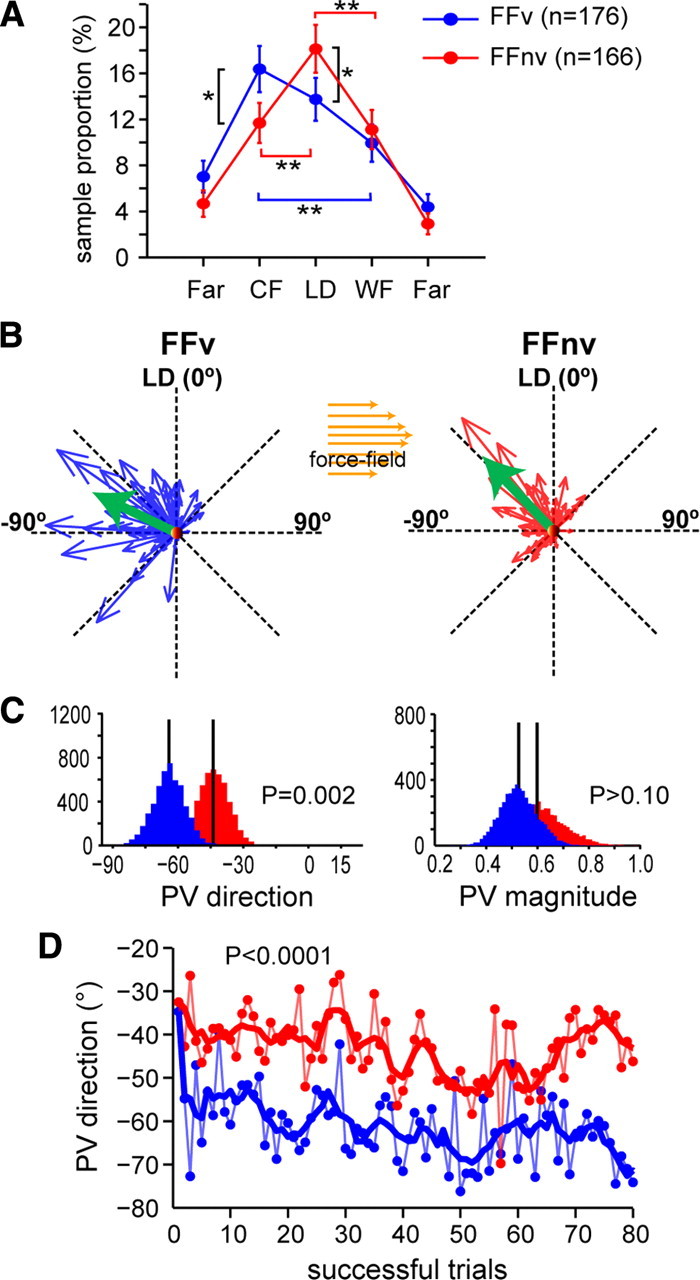
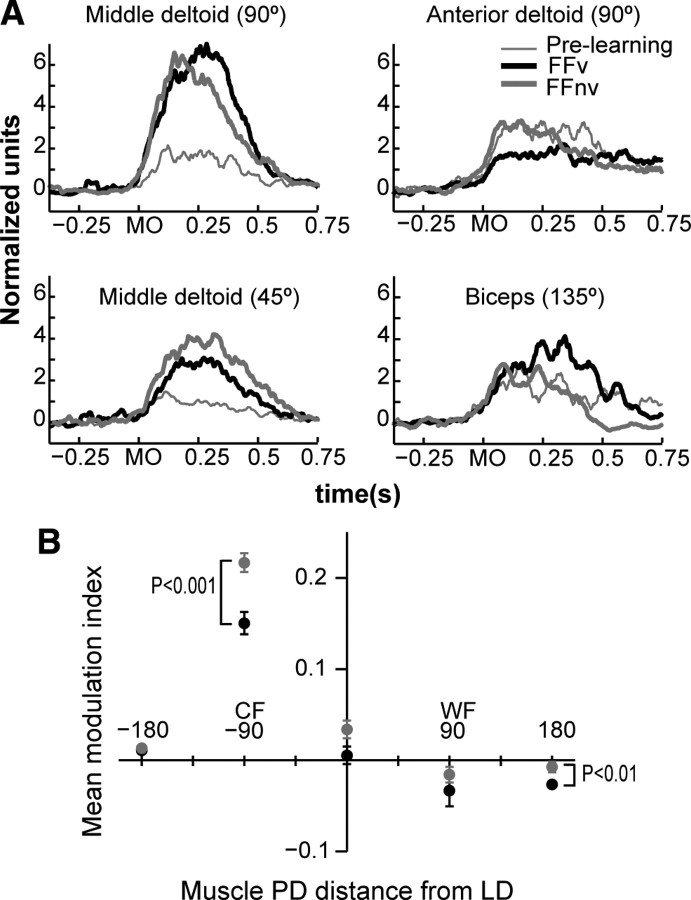
Learning motor skills entails adaptation of neural computations that can generate or modify associations between sensations and actions. Indeed, humans can use different strategies when adapting to dynamic loads depending on available sensory feedback. Here, we examined how neural activity in motor cortex was modified when monkeys made arm reaches to a visual target and locally adapted to curl force field with or without visual trajectory feedback. We found that firing rates of a large subpopulation of cells were consistently modulated depending on the distance of their preferred direction from the learned movement direction. The newly acquired activity followed a cosine-like function, with maximal increase in directions that opposed the perturbing force and decrease in opposite directions. As a result, the combined neuronal activity generated an adapted population vector. The results suggest that this could be achieved without changing the tuning properties of the cells. This population directional signal was however altered in the absence of visual feedback; while the cosine pattern of modulation was maintained, the population distributions of modulated cells differed across feedback consistent with the different trajectory shapes. Finally, we predicted generalization patterns of force-field learning based on the cosine-like modulation. These conformed to reported features of generalization in humans, suggesting that the generalization function was related to the observed rate modulations in the motor cortex. Overall, the findings suggest that the new combined activation of neuronal ensembles could underlie the change in the internal model of movement dynamics in a way that depends on available sensory feedback and chosen strategy.
Figures
References
-
- Arce F, Novick I, Vaadia E. Washington, DC: Society for Neuroscience; 2008. Learning-induced modulation of motor cortical activity during adaptations to force fields with and without visual feedback. Program No. 277.13.
-
- Ashe J, Georgopoulos AP. Movement parameters and neural activity in motor cortex and area 5. Cereb Cortex. 1994;4:590–600. - PubMed
-
- Batschelet E. Circular statistics in biology. London: Academic; 1981.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Research Materials