

Structure of the histone chaperone CIA/ASF1-double bromodomain complex linking histone modifications and site-specific histone eviction
- PMID: 20393127
- PMCID: PMC2889523
- DOI: 10.1073/pnas.0912509107
Structure of the histone chaperone CIA/ASF1-double bromodomain complex linking histone modifications and site-specific histone eviction
Abstract
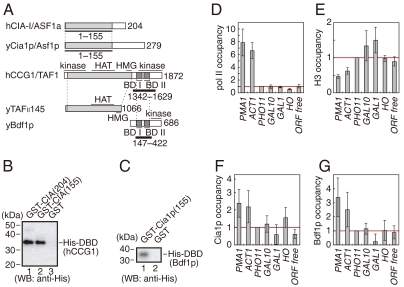
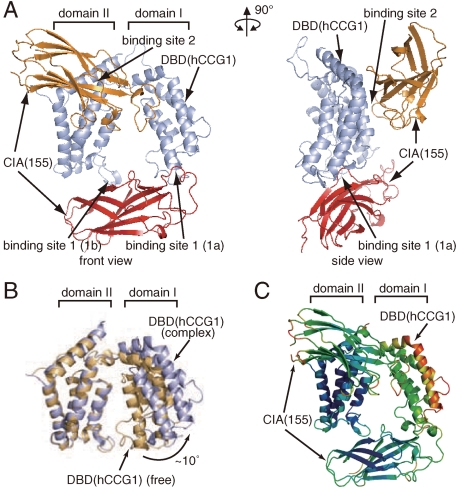
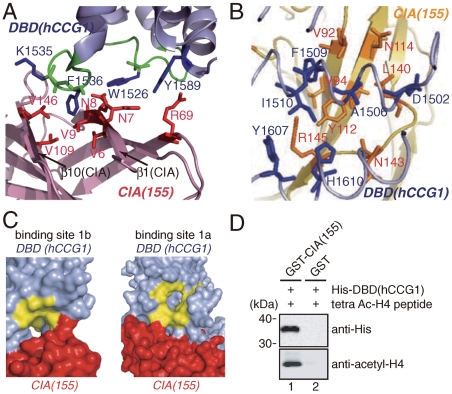
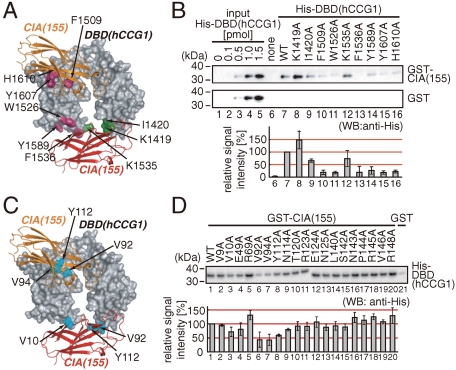
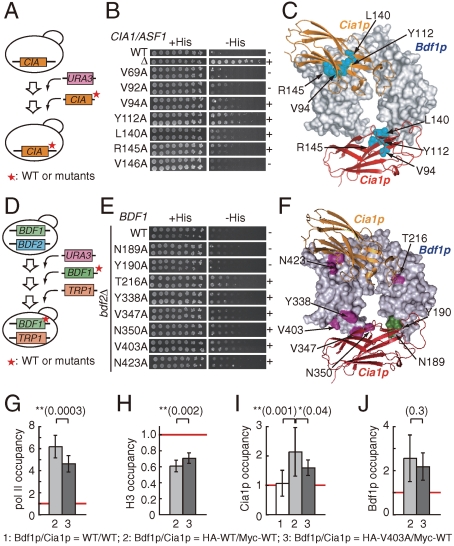
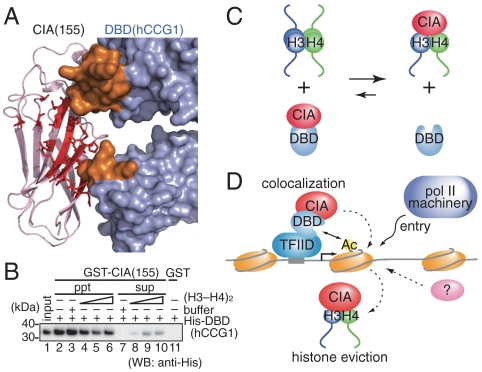
Nucleosomes around the promoter region are disassembled for transcription in response to various signals, such as acetylation and methylation of histones. Although the interactions between histone-acetylation-recognizing bromodomains and factors involved in nucleosome disassembly have been reported, no structural basis connecting histone modifications and nucleosome disassembly has been obtained. Here, we determined at 3.3 A resolution the crystal structure of histone chaperone cell cycle gene 1 (CCG1) interacting factor A/antisilencing function 1 (CIA/ASF1) in complex with the double bromodomain in the CCG1/TAF1/TAF(II)250 subunit of transcription factor IID. Structural, biochemical, and biological studies suggested that interaction between double bromodomain and CIA/ASF1 is required for their colocalization, histone eviction, and pol II entry at active promoter regions. Furthermore, the present crystal structure has characteristics that can connect histone acetylation and CIA/ASF1-mediated histone eviction. These findings suggest that the molecular complex between CIA/ASF1 and the double bromodomain plays a key role in site-specific histone eviction at active promoter regions. The model we propose here is the initial structure-based model of the biological signaling from histone modifications to structural change of the nucleosome (hi-MOST model).
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Kornberg RD. Chromatin structure: A repeating unit of histones and DNA. Science. 1974;184:868–871. - PubMed
-
- Luger K, Mäder AW, Richmond RK, Sargent DF, Richmond TJ. Crystal structure of the nucleosome core particle at 2.8 Å resolution. Nature. 1997;389:251–260. - PubMed
-
- Groth A, Rocha W, Verreault A, Almouzni G. Chromatin challenges during DNA replication and repair. Cell. 2007;128:721–733. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
