

Plastic synaptic networks of the amygdala for the acquisition, expression, and extinction of conditioned fear
- PMID: 20393190
- PMCID: PMC2856122
- DOI: 10.1152/physrev.00037.2009
Plastic synaptic networks of the amygdala for the acquisition, expression, and extinction of conditioned fear
Erratum in
- Physiol Rev. 2010 Jul;90(3):1269
Abstract
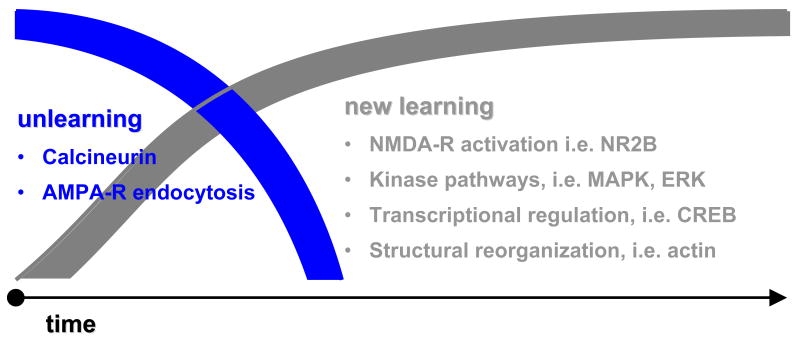
The last 10 years have witnessed a surge of interest for the mechanisms underlying the acquisition and extinction of classically conditioned fear responses. In part, this results from the realization that abnormalities in fear learning mechanisms likely participate in the development and/or maintenance of human anxiety disorders. The simplicity and robustness of this learning paradigm, coupled with the fact that the underlying circuitry is evolutionarily well conserved, make it an ideal model to study the basic biology of memory and identify genetic factors and neuronal systems that regulate the normal and pathological expressions of learned fear. Critical advances have been made in determining how modified neuronal functions upon fear acquisition become stabilized during fear memory consolidation and how these processes are controlled in the course of fear memory extinction. With these advances came the realization that activity in remote neuronal networks must be coordinated for these events to take place. In this paper, we review these mechanisms of coordinated network activity and the molecular cascades leading to enduring fear memory, and allowing for their extinction. We will focus on Pavlovian fear conditioning as a model and the amygdala as a key component for the acquisition and extinction of fear responses.
Figures
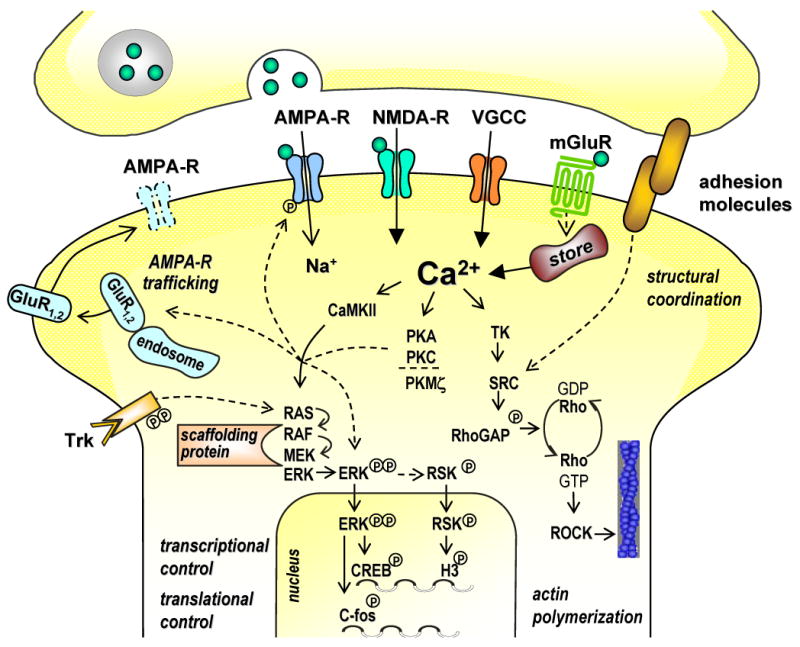
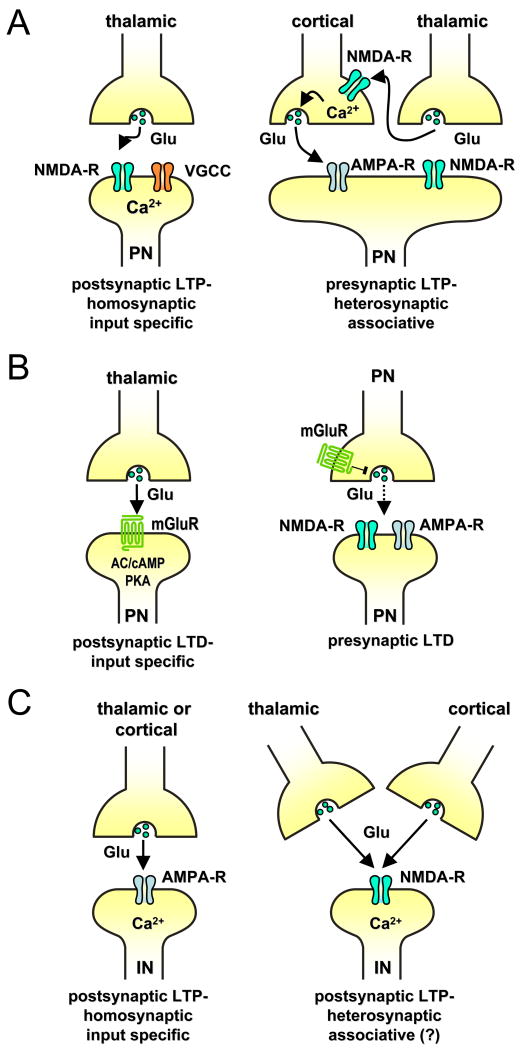
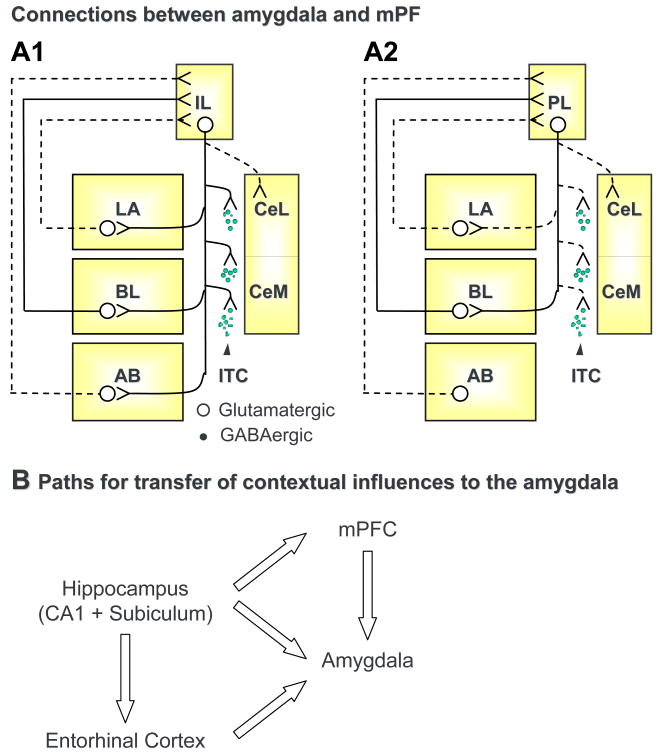
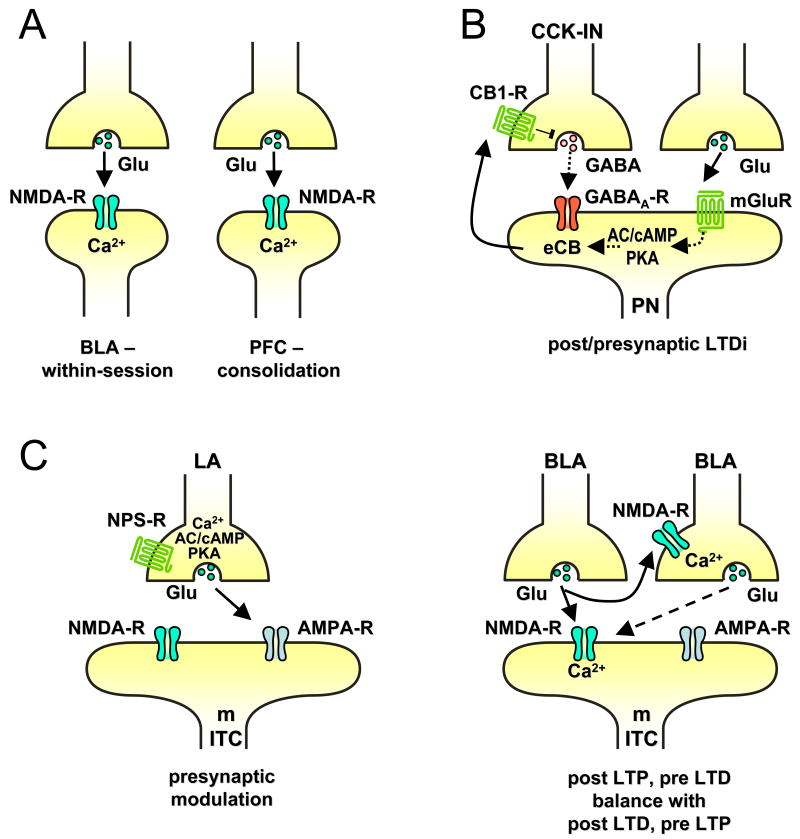
References
-
- Abraham WC. Metaplasticity: tuning synapses and networks for plasticity. Nat Rev Neurosci. 2008;9:387. - PubMed
-
- Aggleton JP, Saunders RC. The amygdala - What's happened in the last decade? In: Aggleton JP, editor. The Amygdala. Oxford, UK: Oxford University Press; 2000. pp. 1–30.
-
- Aitkin LM, Irvine DR, Nelson JE, Merzenich MM, Clarey JC. Frequency representation in the auditory midbrain and forebrain of a marsupial, the northern native cat (Dasyurus hallucatus) Brain Behav Evol. 1986;29:17–28. - PubMed
-
- Albrecht A, Bergado J, Pape HC, Stork O. Role of NCAM in amygdalo-hippocampal interactions and stress modulation of context fear memory in revision. 2009.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical