

A novel native store-operated calcium channel encoded by Orai3: selective requirement of Orai3 versus Orai1 in estrogen receptor-positive versus estrogen receptor-negative breast cancer cells
- PMID: 20395295
- PMCID: PMC2885196
- DOI: 10.1074/jbc.M110.102582
A novel native store-operated calcium channel encoded by Orai3: selective requirement of Orai3 versus Orai1 in estrogen receptor-positive versus estrogen receptor-negative breast cancer cells
Abstract
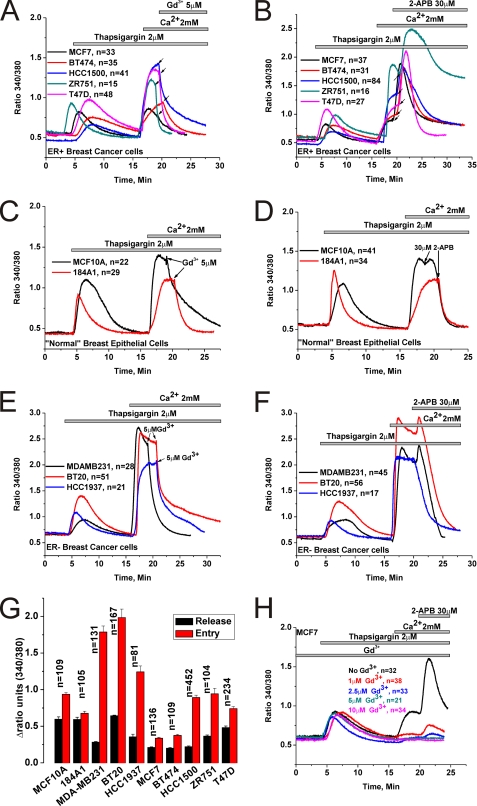
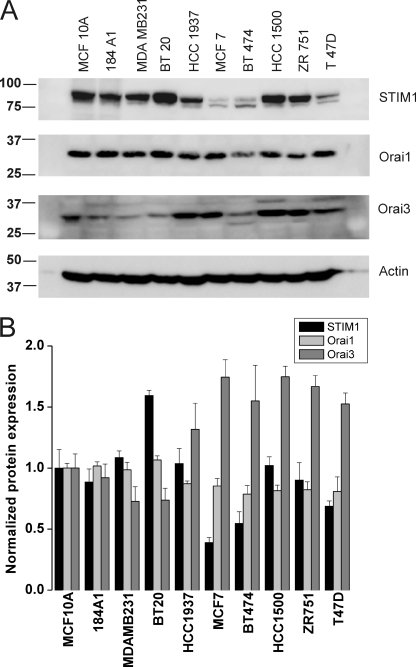
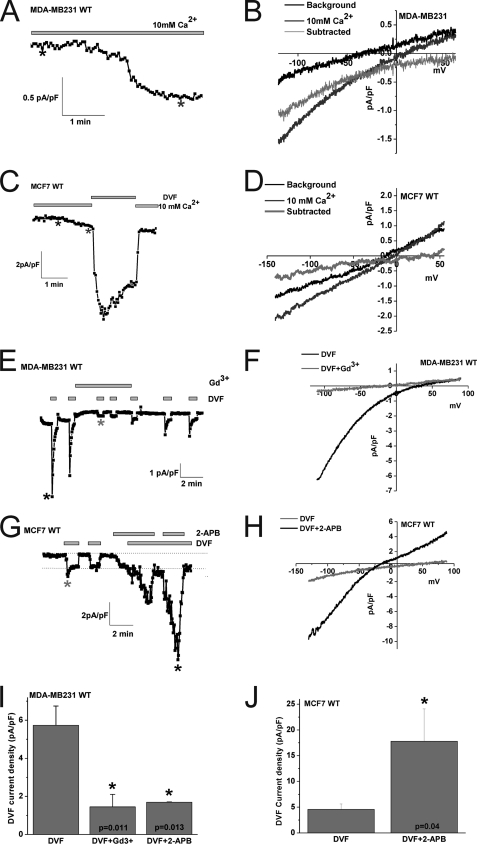
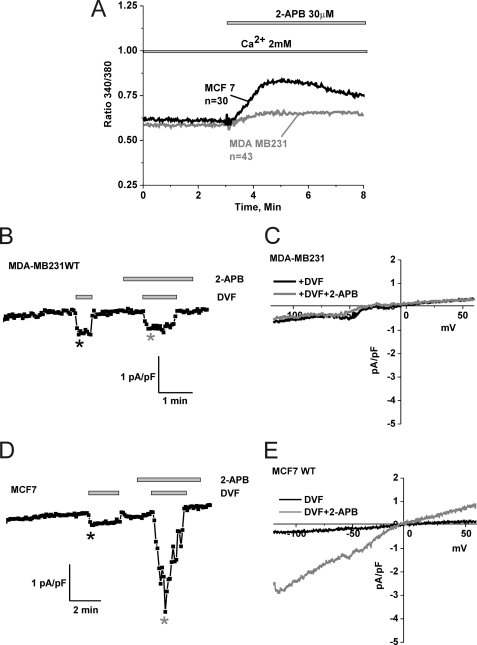
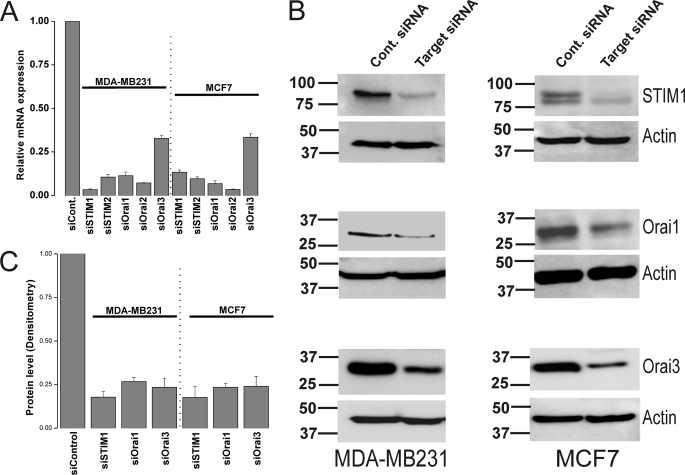
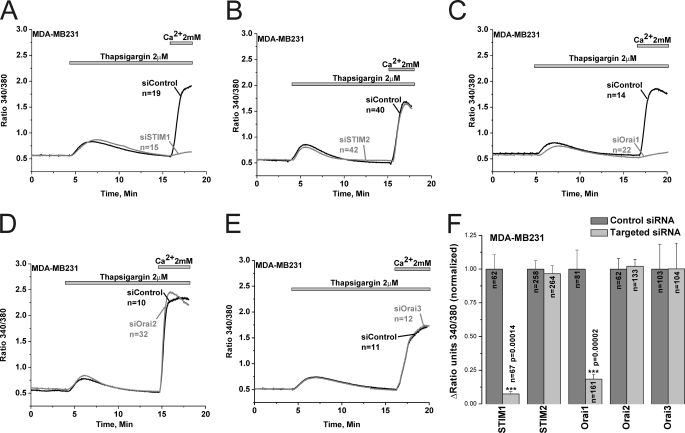
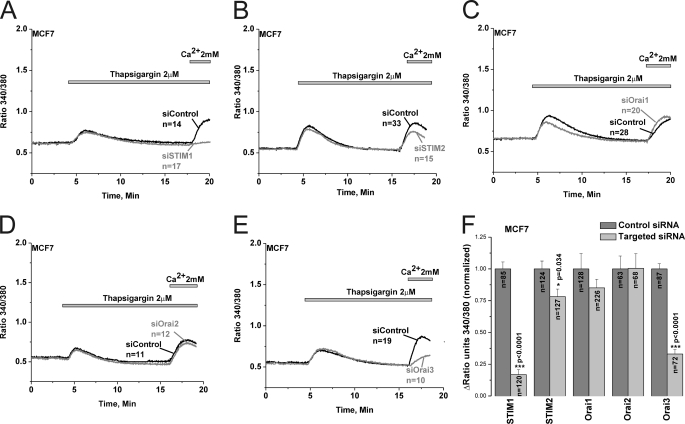
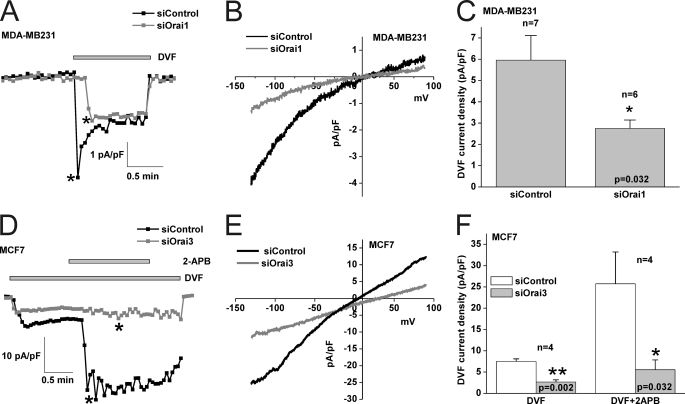
Store-operated calcium (Ca(2+)) entry (SOCE) mediated by STIM/Orai proteins is a ubiquitous pathway that controls many important cell functions including proliferation and migration. STIM proteins are Ca(2+) sensors in the endoplasmic reticulum and Orai proteins are channels expressed at the plasma membrane. The fall in endoplasmic reticulum Ca(2+) causes translocation of STIM1 to subplasmalemmal puncta where they activate Orai1 channels that mediate the highly Ca(2+)-selective Ca(2+) release-activated Ca(2+) current (I(CRAC)). Whereas Orai1 has been clearly shown to encode SOCE channels in many cell types, the role of Orai2 and Orai3 in native SOCE pathways remains elusive. Here we analyzed SOCE in ten breast cell lines picked in an unbiased way. We used a combination of Ca(2+) imaging, pharmacology, patch clamp electrophysiology, and molecular knockdown to show that native SOCE and I(CRAC) in estrogen receptor-positive (ER(+)) breast cancer cell lines are mediated by STIM1/2 and Orai3 while estrogen receptor-negative (ER(-)) breast cancer cells use the canonical STIM1/Orai1 pathway. The ER(+) breast cancer cells represent the first example where the native SOCE pathway and I(CRAC) are mediated by Orai3. Future studies implicating Orai3 in ER(+) breast cancer progression might establish Orai3 as a selective target in therapy of ER(+) breast tumors.
Figures
References
-
- Glass A. G., Lacey J. V., Jr., Carreon J. D., Hoover R. N. (2007) J. Natl. Cancer Inst. 99, 1152–1161 - PubMed
-
- Bai Z., Gust R. (2009) Archiv der Pharmazie 342, 133–149 - PubMed
-
- Hurvitz S. A., Finn R. S. (2009) Future Oncology 5, 1015–1025 - PubMed
-
- Parekh A. B., Putney J. W., Jr. (2005) Physiol. Rev. 85, 757–810 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
