

A Cladistic Approach for the Classification of Oligotrichid Ciliates (Ciliophora: Spirotricha)
- PMID: 20396404
- PMCID: PMC2854820
A Cladistic Approach for the Classification of Oligotrichid Ciliates (Ciliophora: Spirotricha)
Abstract
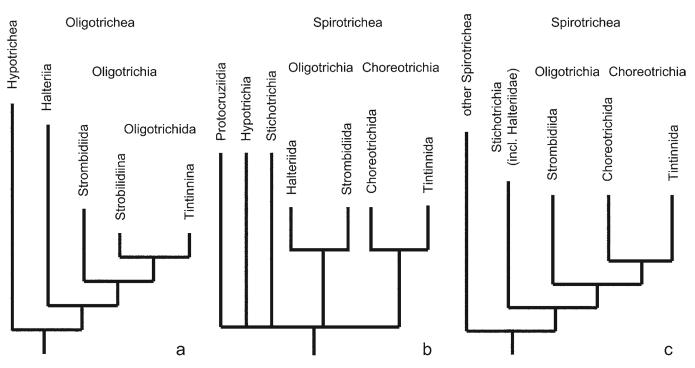
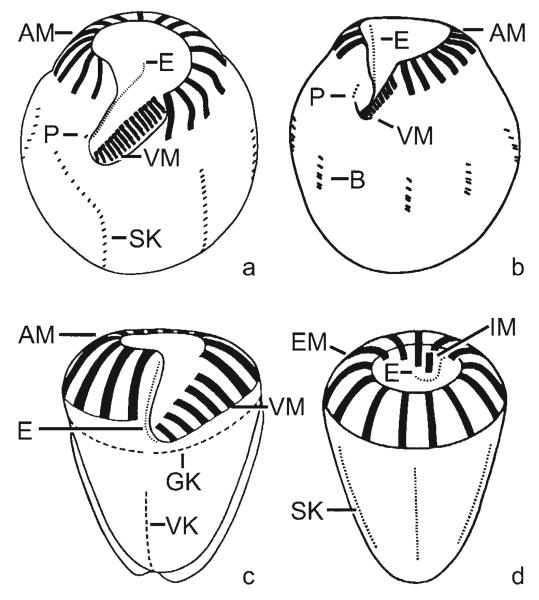
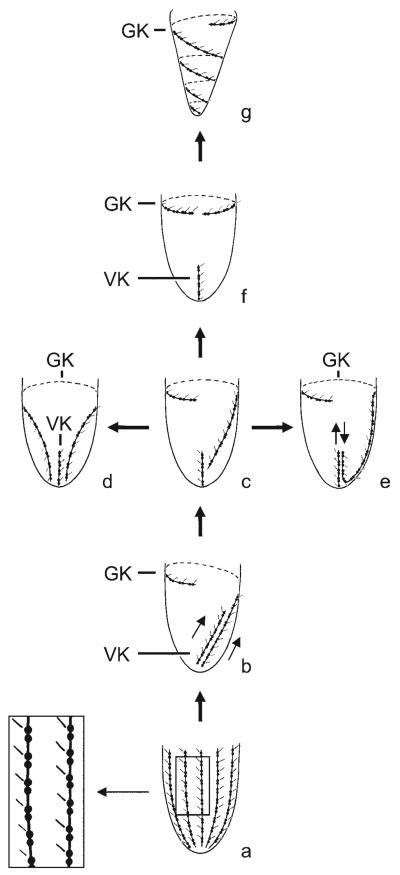
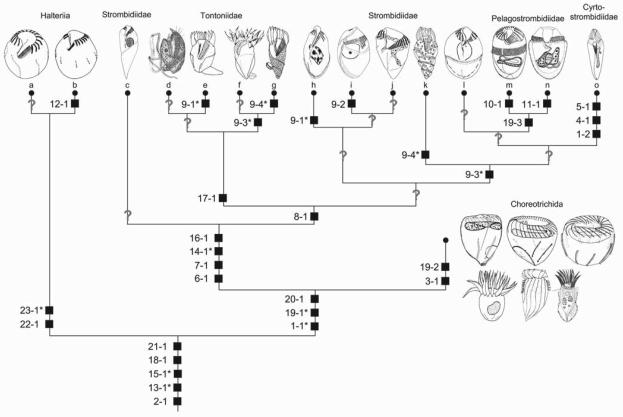
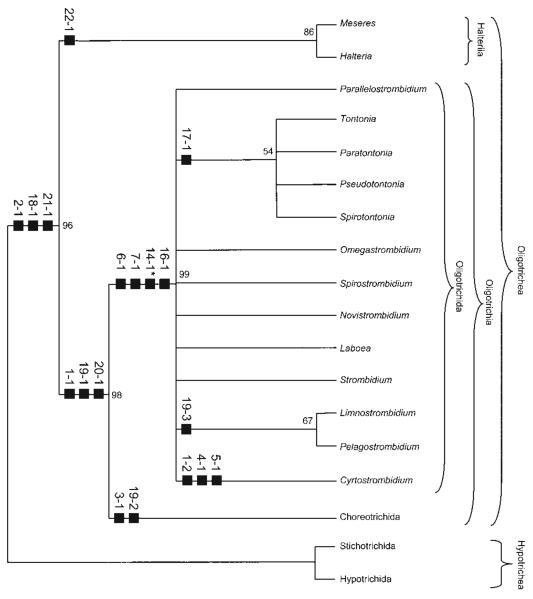
Currently, gene sequence genealogies of the Oligotrichea Bütschli, 1889 comprise only few species. Therefore, a cladistic approach, especially to the Oligotrichida, was made, applying Hennig's method and computer programs. Twenty-three characters were selected and discussed, i.e., the morphology of the oral apparatus (five characters), the somatic ciliature (eight characters), special organelles (four characters), and ontogenetic particulars (six characters). Nine of these characters developed convergently twice. Although several new features were included into the analyses, the cladograms match other morphological trees in the monophyly of the Oligotrichea, Halteriia, Oligotrichia, Oligotrichida, and Choreotrichida. The main synapomorphies of the Oligotrichea are the enantiotropic division mode and the de novo-origin of the undulating membranes. Although the sister group relationship of the Halteriia and the Oligotrichia contradicts results obtained by gene sequence analyses, no morphologic, ontogenetic or ultrastructural features were found, which support a branching of Halteria grandinella within the Stichotrichida. The cladistic approaches suggest paraphyly of the family Strombidiidae probably due to the scarce knowledge. A revised classification of the Oligotrichea is suggested, including all sufficiently known families and genera.
Figures
References
-
- Aescht E. Catalogue of the generic names of ciliates (Protozoa, Ciliophora) Denisia. 2001;1
-
- Agatha S. Morphology and ontogenesis of Novistrombidium apsheronicum nov. comb. and Strombidium arenicola (Protozoa, Ciliophora): a comparative light microscopical and SEM study. Europ. J. Protistol. 2003a;39:245–266.
-
- Agatha S. Redescription of Strombidinopsis minima (Gruber, 1884) Lynn et al., 1991 (Protozoa, Ciliophora), with notes on its ontogenesis and distribution. Europ. J. Protistol. 2003b;39:233–244.
-
- Agatha S, Riedel-Lorjé JC. Morphology, infraciliature, and ecology of halteriids and strombidiids (Ciliophora, Oligotrichea) from coastal brackish water basins. Arch. Protistenk. 1997;148:445–459.
Grants and funding
LinkOut - more resources
Full Text Sources