

Picornaviruses
- PMID: 20397067
- PMCID: PMC3018333
- DOI: 10.1007/82_2010_37
Picornaviruses
Abstract
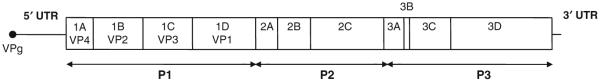
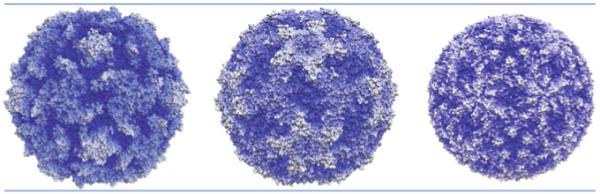
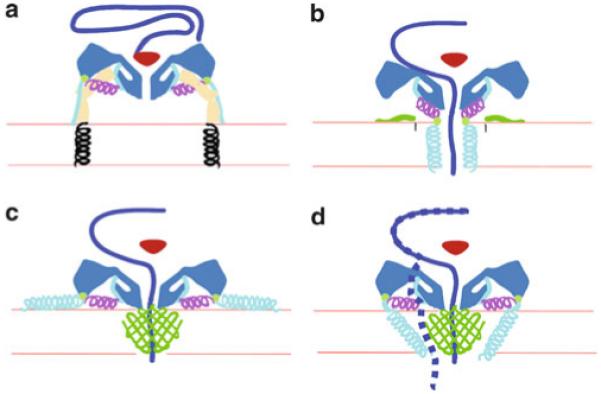
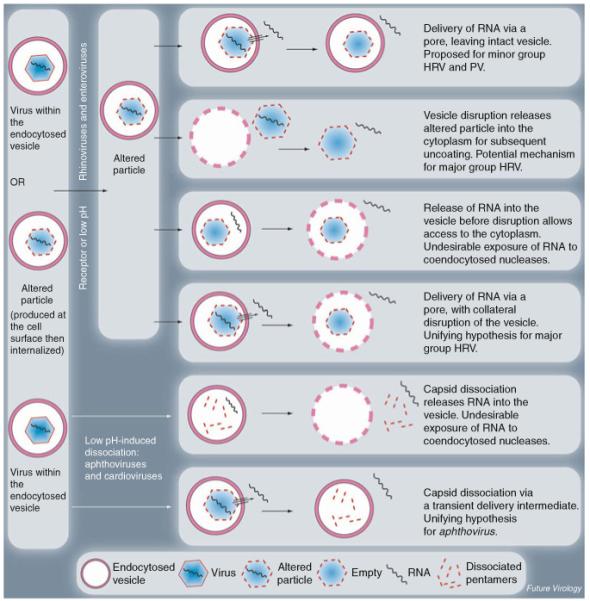
The picornavirus family consists of a large number of small RNA viruses, many of which are significant pathogens of humans and livestock. They are amongst the simplest of vertebrate viruses comprising a single stranded positive sense RNA genome within a T = 1 (quasi T = 3) icosahedral protein capsid of approximately 30 nm diameter. The structures of a number of picornaviruses have been determined at close to atomic resolution by X-ray crystallography. The structures of cell entry intermediate particles and complexes of virus particles with receptor molecules or antibodies have also been obtained by X-ray crystallography or at a lower resolution by cryo-electron microscopy. Many of the receptors used by different picornaviruses have been identified, and it is becoming increasingly apparent that many use co-receptors and alternative receptors to bind to and infect cells. However, the mechanisms by which these viruses release their genomes and transport them across a cellular membrane to gain access to the cytoplasm are still poorly understood. Indeed, detailed studies of cell entry mechanisms have been made only on a few members of the family, and it is yet to be established how broadly the results of these are applicable across the full spectrum of picornaviruses. Working models of the cell entry process are being developed for the best studied picornaviruses, the enteroviruses. These viruses maintain particle integrity throughout the infection process and function as genome delivery modules. However, there is currently no model to explain how viruses such as cardio- and aphthoviruses that appear to simply dissociate into subunits during uncoating deliver their genomes into the cytoplasm.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
- G0200504/MRC_/Medical Research Council/United Kingdom
- BB/E00931X/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BBS/E/I/00001411/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- G0600025/MRC_/Medical Research Council/United Kingdom
- R01 AI020566/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
