

Translocation of Borrelia burgdorferi surface lipoprotein OspA through the outer membrane requires an unfolded conformation and can initiate at the C-terminus
- PMID: 20398211
- PMCID: PMC2999405
- DOI: 10.1111/j.1365-2958.2010.07172.x
Translocation of Borrelia burgdorferi surface lipoprotein OspA through the outer membrane requires an unfolded conformation and can initiate at the C-terminus
Abstract
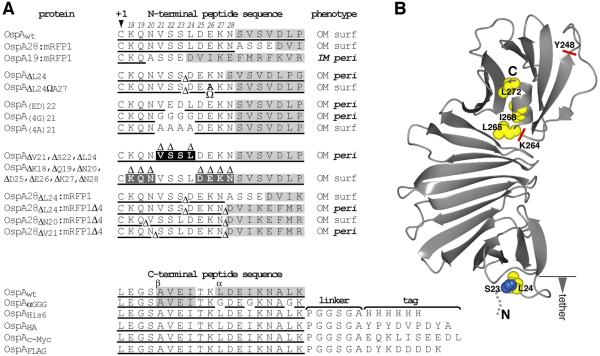
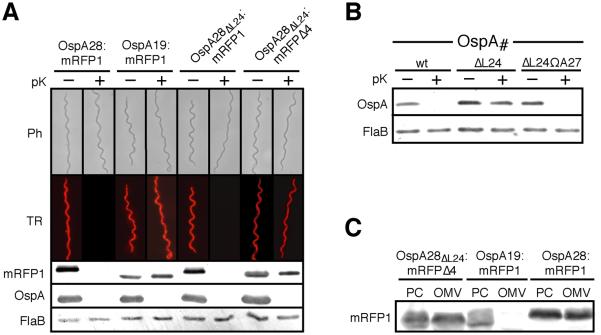
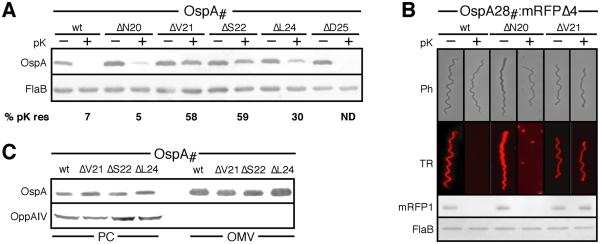
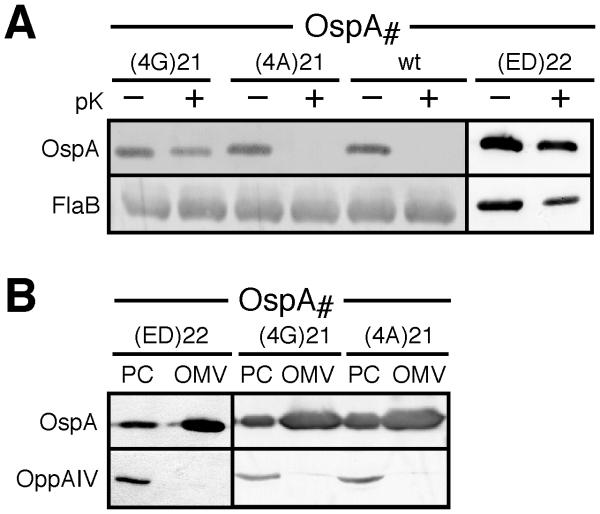
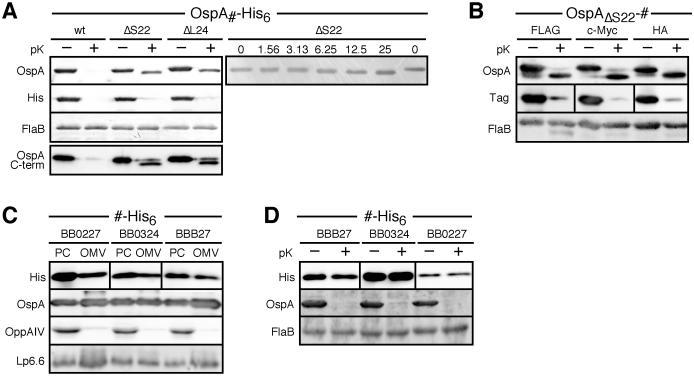
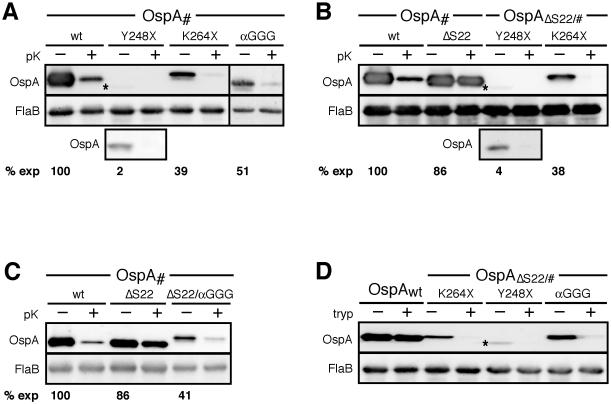
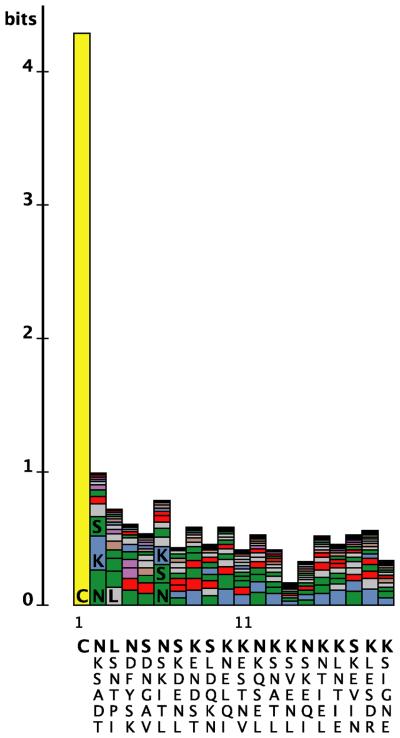
Borrelia burgdorferi surface lipoproteins are essential to the pathogenesis of Lyme borreliosis, but the mechanisms responsible for their localization are only beginning to emerge. We have previously demonstrated the critical nature of the amino-terminal 'tether' domain of the mature lipoprotein for sorting a fluorescent reporter to the Borrelia cell surface. Here, we show that individual deletion of four contiguous residues within the tether of major surface lipoprotein OspA results in its inefficient translocation across the Borrelia outer membrane. Intriguingly, C-terminal epitope tags of these N-terminal deletion mutants were selectively surface-exposed. Fold-destabilizing C-terminal point mutations and deletions did not block OspA secretion, but rather restored one of the otherwise periplasmic tether mutants to the bacterial surface. Together, these data indicate that disturbance of a confined tether feature leads to premature folding of OspA in the periplasm and thereby prevents secretion through the outer membrane. Furthermore, they suggest that OspA emerges tail-first on the bacterial surface, yet independent of a specific C-terminal targeting peptide sequence.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
